Open Access | Review
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Role of miR-140-5p in vascular aging and arterial stiffening: a review of potential mechanisms
* Corresponding author: Muhammad Iqhrammullah
Mailing address: Postgraduate Program of Public Health, Universitas Muhammadiyah Aceh, Banda Aceh, Indonesia.
Email: m.iqhrammullah@gmail.com
Received: 24 March 2026 / Revised: 28 April 2026 / Accepted: 11 May 2026 / Published: 30 June 2026
DOI: 10.31491/APT.2026.06.217
Abstract
Vascular aging is characterized by progressive endothelial dysfunction, arterial stiffening, and structural remodeling of the vascular wall, which are processes that precede overt cardiovascular disease and predict adverse outcomes independently. MicroRNAs have emerged as key post-transcriptional regulators linking cellular aging mechanisms to clinically measurable vascular phenotypes and their attenuation. Among them, miR-140-5p has gained attention for its role in maintaining vascular homeostasis and modulating aging-related vascular dysfunction. Experimental evidence indicates that miR-140-5p expression decreases with age and vascular stress due to epigenetic modifications, including promoter hyper-methylation and reduced miRNA biogenesis capacity, contributing to endothelial senescence, senescence-associated secretory phenotype (SASP), impaired nitric oxide bioavailability, and chronic low-grade inflammation. At the medial layer, reduced miR-140-5p favors vascular smooth muscle cell phenotypic switching and extracellular matrix imbalance, promoting arterial stiffening. These molecular alterations provide a mechanistic basis linking miR-140-5p dysregulation to established clinical indices of vascular aging, including reduced flow-mediated dilation and increased pulse wave velocity. This review synthesizes current evidence on the biological role of miR-140-5p in vascular aging, integrates microRNA molecular mechanisms with functional and structural vascular metrics, and discusses its potential as a biomarker and therapeutic target. Understanding the role of miR-140-5p may support strategies aimed at preserving vascular resilience and delaying age-related cardiovascular risk.
Keywords
MicroRNA, vascular aging, endothelial dysfunction, arterial stiffness, pulse wave velocity
Introduction
Vascular aging is a progressive and multifactorial process characterized by endothelial dysfunction, arterial stiffening, and structural remodeling of the vascular wall [1, 2]. These changes occur gradually with advancing age and precede the development of overt cardiovascular disease, independently leading to increased cardiovascular morbidity and mortality even in the absence of traditional risk factors [2, 3]. Hallmark features of vascular aging include impaired endothelium-dependent vasodilation, increased collagen deposition with elastin fragmentation, vascular smooth muscle cell (VSMC) phenotypic switching, and chronic low-grade inflammation, collectively leading to reduced arterial compliance and elevated pulsatile load [2, 3]. Clinically, vascular aging is commonly quantified using functional and structural indices. Flow-mediated dilation (FMD) serves as a noninvasive surrogate of endothelial nitric oxide (NO)-dependent vasodilatory capacity and reflects early, potentially reversible vascular dysfunction [4]. Carotid-femoral pulse wave velocity (PWV), the gold-standard measure of arterial stiffness, integrates cumulative functional impairment and structural remodeling of the arterial wall and is a strong predictor of cardiovascular events and all-cause mortality [5]. Importantly, deterioration in FMD often precedes measurable increases in PWV, supporting the concept that endothelial dysfunction represents an early initiating event in the vascular aging continuum [5].
In recent years, increasing attention has focused on the molecular mechanisms that link cellular aging to these clinically measurable vascular phenotypes [1, 2]. Among these, microRNAs have emerged as key post-transcriptional regulators of vascular homeostasis, modulating endothelial function, VSMC behavior, extracellular matrix (ECM) turnover, and inflammatory signaling [6, 7]. Age-associated alterations in microRNA expression profiles have been consistently observed in vascular tissues and circulating compartments, suggesting that dysregulated microRNA networks contribute to the initiation and progression of vascular aging rather than merely reflecting downstream disease states [8, 9]. In this light, miR-140-5p has gained interest as a biologically plausible regulator of vascular aging. Although initially characterized in musculoskeletal tissues, accumulating evidence indicates that miR-140-5p is expressed in endothelial cells and VSMCs and responds to aging-related stressors such as oxidative stress, inflammation, and metabolic perturbations [10-12]. Experimental studies suggest that miR-140-5p participates in pathways relevant to cellular senescence, NO bioavailability, inflammatory activation, and ECM regulation—processes central to endothelial dysfunction and arterial stiffening [10]. Importantly, miR-140-5p expression has been suggested to decline with age in animal models [10, 13], positioning it as a potential upstream modulator linking molecular aging mechanisms to functional and structural vascular alterations. This review synthesizes current evidence on the role of miR-140-5p in vascular aging, with a particular focus on its mechanistic connections to endothelial dysfunction, arterial stiffness, and clinically relevant indices such as FMD and PWV. By integrating molecular insights with established vascular aging metrics, the work aims to clarify the relevance of miR-140-5p within the broader framework of cardiovascular aging.
Methods
This narrative review was done through literature search from PubMed, Scopus, Cochrane, and ScienceDirect databases using the keywords "miR-140-5p", "vascular aging", and "arterial stiffness". We included studies which assessed potential mechanisms and the role of miR-140-5p in vascular aging and arterial stiffening, along with related literature.
Biological profile of miR-140-5p
miR-140-5p is the predominant mature strand derived from the miR-140 precursor and has been increasingly recognized for its regulatory roles in vascular and aging-related processes [14]. The miR-140 locus generates both miR-140-5p and miR-140-3p from the same precursor;
however, miR-140-5p is generally the more abundant and functionally relevant form in vascular cells [10]. To date, miR-140-5p has been most extensively studied in musculoskeletal tissues, particularly in cartilage, where it is a key regulator of chondrocyte homeostasis and is consistently implicated in age-related disorders such as osteoarthritis and rheumatoid arthritis [6, 14, 15]. This established role in tissue aging and ECM turnover provides a biological basis for investigating its function in the vasculature. It is expressed in endothelial cells and VSMCs, where it participates in post-transcriptional regulation of genes involved in cell proliferation, apoptosis, ECM remodeling, and inflammatory signaling [16]. Importantly, miR-140-5p expression is modulated by cellular stressors that are highly relevant to vascular aging, including oxidative stress, inflammation, and metabolic perturbations which characterize age-related vascular dysfunction [10].
Emerging evidence indicates that miR-140-5p expression changes dynamically in response to pathological stimuli. For example, in models of atherosclerosis and oxidative injury, miR-140-5p modulates redox homeostasis by interacting with the Nrf2/Sirt2/Keap1 antioxidant pathway, exacerbating oxidative stress when overexpressed and contributing to vascular dysfunction [17]. Another study in Hutchinson-Gilford progeria syndrome (HGPS) fibroblasts—models of accelerated aging—demonstrated that miR-140-5p overexpression impairs nuclear factor erythroid 2-related factor 2 (Nrf2) signaling, increases oxidative stress, and induces mitochondrial dysfunction, further implicating miR-140-5p in aging-associated cellular stress responses [18].
While the bulk of vascular research on miR-140-5p predates 2024, mechanistic studies consistently show that miR-140-5p regulates processes central to vascular cell homeostasis. In human VSMCs exposed to oxidized low-density lipoprotein (ox-LDL), miR-140-5p influences viability, migration, and apoptosis via interaction with targets such as ROBO4, highlighting its involvement in atherogenic cellular behavior [19]. Additionally, miR-140-5p and related regulatory networks are implicated across aging-related contexts, interacting with long non-coding RNAs and other molecular regulators that influence cellular senescence, inflammation, and ECM metabolism [10].
Epigenetic and transcriptional regulation of miR-140-5p in vascular aging
It is noteworthy that the age-related decrease in miR-140-5p expression does not occur passively but is mediated by active changes at the epigenetic and transcriptional levels. Understanding the upstream regulatory mechanisms is necessary to identify potential interventions that could restore miR-140-5p levels in aging vascular tissue. The miR-140 gene is located on chromosome 16q22.1 within intron 15 of the WWP2 gene (WW domain containing E3 ubiquitin protein ligase 2), so its expression depends on host gene regulation [10, 20]. Epigenomic analysis revealed a differentially methylated region (DMR) spanning 228 base pairs with 16 CpG sites in the WWP2 promoter that controls miR-140 transcription [21].
Studies using dCas9-DNMT3a epigenetic editing demonstrated that experimental hypermethylation of this DMR simultaneously decreased WWP2 and miR-140 expression, indicating a fairly strong causal relationship [21]. In aging cartilage tissue, increased CpG methylation in the miR-140-5p regulatory region is correlated with decreased
expression and reduced binding affinity of the SMAD3 transcription factor [22]. This evidence was confirmed by the administration of 5-aza-2′-deoxycytidine as a DNA demethylating agent, which successfully restored miR-140-5p
expression in hypermethylated chondrocytes [22].
Methylation data on aged endothelial and VSMCs are still limited; however, global DNA methylation changes have
been recorded in both cell types during vascular aging, suggesting that similar mechanisms may
occur [23]. The transcriptional regulation of miR-140
is influenced by transcription factors that respond to oxidative stress and inflammation. Nrf2
directly binds the miR-140 promoter region and enhances its transcription, as validated by a
chromatin immunoprecipitation (ChIP) assay. Knockdown of Nrf2 via small interfering RNA (siRNA)
significantly decreased miR-140 expression, confirming the role of Nrf2 as a transcriptional
activator [24]. Conversely, the activation of NF-κB
under chronic inflammatory conditions suppresses miR-140-5p transcription through competition
with transcriptional activators in the promoter region [25].
This forms a pathological feedback loop of oxidative stress and inflammation that reduces miR-140-5p,
thereby exacerbating redox dysfunction and the inflammatory response, both of which are hallmarks of
vascular aging [26]. The maturation of miR-140-5p from
its precursor into its functional form requires the activity of microRNA-processing enzymes, whose
capacity is reduced in aging cells. The maturation process involves the cleavage of pri-miRNA by
the Drosha-DGCR8 complex in the nucleus and the cleavage of pre-miRNA by Dicer in the cytoplasm.
In aged mouse cerebral microvascular endothelial cells, Dicer expression is significantly reduced,
accompanied by a global decrease in mature microRNA profiles and impaired angiogenic capacity. The
overexpression of Dicer in aged endothelial cells partially restores microRNA profiles and angiogenic
function and plays a role in the decline in Dicer in age-related vascular dysfunction
[13, 27].
Genetic deletion of Dicer in endothelial cells alters the expression of specific microRNAs and
affects the vascular inflammatory response [28].
In VSMCs, Drosha knock-down reduces cell viability and proliferation, indicating an important
role for microRNA biogenesis in homeostasis [29].
To date, changes in microRNA processing enzymes have not been adequately characterized for miR-140-5p
in aging vascular cells, so a global reduction in microRNA processing capacity provides a plausible
mechanism for the reduction in mature miR-140-5p.
The regulation of miR-140-5p becomes more complex
in the presence of competing endogenous RNA (ceRNAs), which absorb miR-140-5p and reduce its
availability to suppress target mRNAs. The long noncoding RNA (lncRNA) H19 binds to
miR-140-5p directly in VSMCs, as validated through a luciferase reporter assay and RNA
immunoprecipitation. The overexpression of H19 enhances the osteoblastic transition of
VSMCs by sequestering miR-140-5p and suppressing Satb2 and pro-calcification transcription
factors [30]. Similarly, the lncRNA PCA3 functions as
a ceRNA for miR-140-5p in macrophages, forming the PCA3/miR-140-5p/RFX7/ABCA1 axis, which modulates
lipid accumulation and atherosclerosis progression [31].
Changes in lncRNA expression during vascular aging can shift the balance of miR-140-5p
between its free active form and its ceRNA-bound form, thereby affecting the regulatory capacity of miR-140-5p for its target mRNAs. This layered regulatory mechanism suggests that the decrease in miR-140-5p during vascular aging is the result of the integration of epigenetic changes, transcription factor activity, microRNA biogenesis capacity, and ceRNA networks. This understanding opens up opportunities for multilevel interventions, such as promoter demethylation, modulation of the Nrf2/NF-κB pathway, restoration of Dicer activity, or ceRNA manipulation, which could be strategies to restore miR-140-5p function in aging blood vessels (Table 1).
Table 1.
Summary of miR-140-5p mechanisms in vascular and nonvascular cells.
| Mechanism | Regulation | Effects induced | Cell type | Validation method | Reference |
|---|---|---|---|---|---|
| DNA methylation | CpG sites on the WWP2 promoter (16 sites, 228 bp DMR) | Hypermethylation → ↓ expression | Chondrocytes | dCas9-DNMT3a editing | [21] |
| DNA methylation | miR-140-5p promoter region | Hypermethylation → ↓ SMAD3 binding | Chondrocytes | Bisulfite sequencing, ChIP | [22] |
| Transcription factor | Nrf2 binding | Nrf2 activation → ↑ expression | Pulmonary fibroblasts | ChIP assay, siRNA knockdown | [24] |
| Transcription factor | NF-κB activation | NF-κB activation → ↓ expression | Endothelial cells | qRT-PCR | [25] |
| miRNA biogenesis | Dicer expression | Decreased Dicer → ↓ miR-140-5p maturation | Microvascular endothelial cells | Overexpression rescue | [27] |
| miRNA biogenesis | Drosha-DGCR8 | Knockdown → ↓ VSMC viability | VSMC | siRNA knockdown | [32] |
| ceRNA network | lncRNA H19 | H19 sponging → ↓ bioavailability | VSMC | Luciferase, RIP assay | [30] |
| ceRNA network | lncRNA PCA3 | PCA3 sponging → ↓ bioavailability | Macrophage | Luciferase, RIP assay | [31] |
Note: DMR: differentially methylated region; ChIP: chromatin immunoprecipitation; RIP: RNA immunoprecipitation; VSMC: vascular smooth muscle cell.
miR-140-5p and endothelial function
Endothelial dysfunction is an early and defining feature of vascular aging, often characterized by impaired NO-mediated vasodilation,
heightened oxidative stress, and a pro-inflammatory endothelial phenotype [33].
Endothelial nitric oxide synthase (eNOS)-derived NO is critical for vasodilatory capacity and
vascular homeostasis; reductions in NO bioavailability are strongly associated with diminished
FMD and heightened cardiovascular risk [4, 34].
In light of miRNAs as pivotal regulators of endothelial gene networks [35],
several lines of evidence suggest that miR-140-5p participates in endothelial responses to
metabolic and oxidative stress. In human endothelial cell models exposed to high glucose and
oxidative stimuli, alterations in miR-140-5p expression have been
observed [36]. In a study, miR-140-5p has been implicated
in pathways affecting cellular proliferation, apoptosis, and angiogenic signaling in hyperglycemic
human umbilical vein endothelial cells (HUVECs) [36].
Studies of oxidative stress in endothelial contexts also identify miR-140-5p among miRNAs that influence
redox regulatory networks, including Nrf2-SIRT2 signaling
axes [24, 26].
Overexpression of miR-140-5p under oxidative conditions has been shown to increase reactive oxygen
species (ROS) production and reduce antioxidant activity in HUVECs exposed to ox-LDL, suggesting
that dysregulated miR-140-5p may exacerbate redox imbalance in endothelial
cells [37]. This aligns with broader evidence that
miRNAs can modulate endothelial senescence and inflammation by targeting redox and NF-κB pathways
that converge on NO signaling and vasodilatory function [35].
Furthermore, an epigenetic study suggests that downregulation of miR-140-5p in the aorta may be linked
to atherosclerosis by influencing endothelial gene expression (Figure 1) [38].
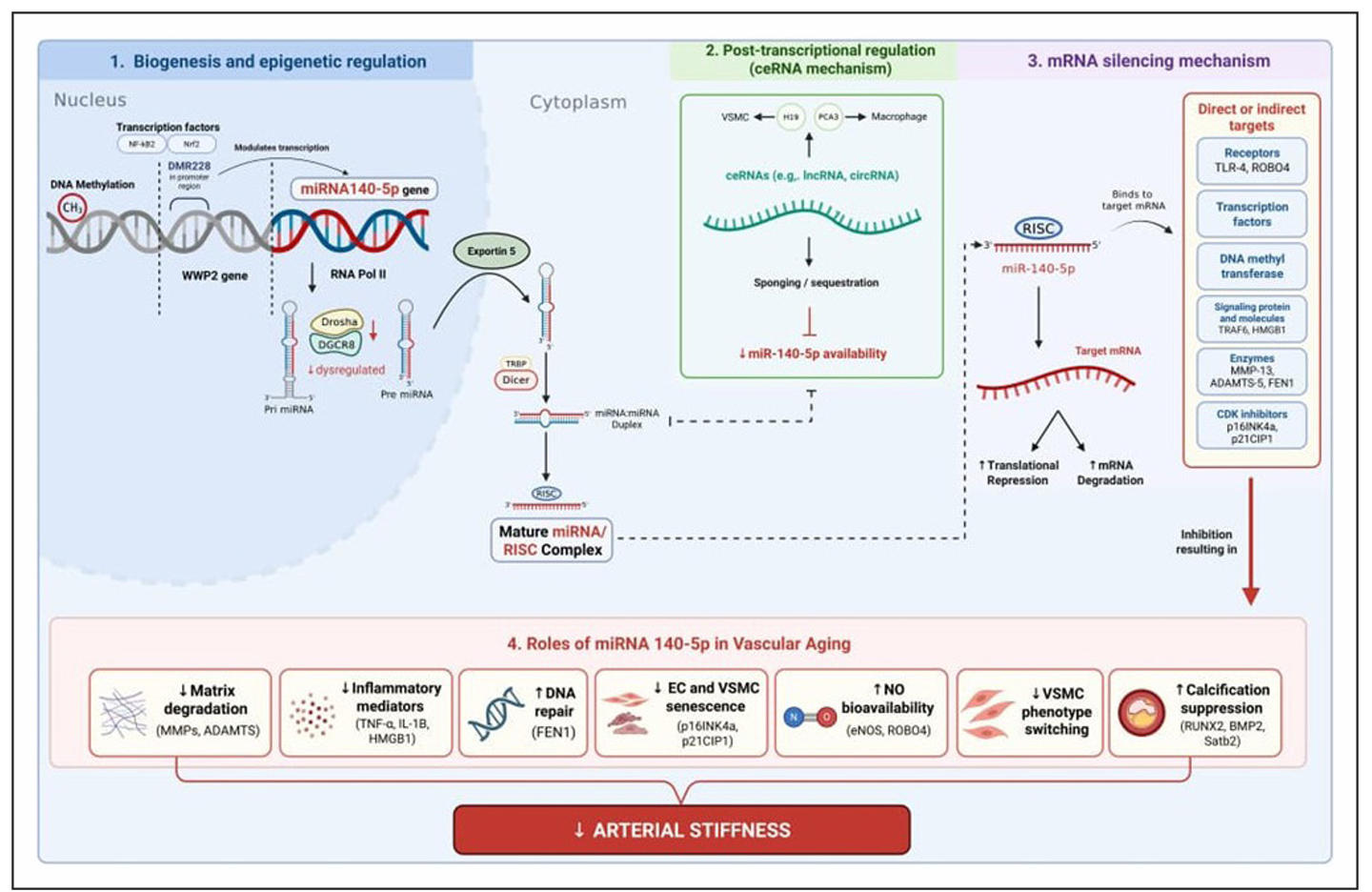
Figure 1. Mechanisms, pathway, and roles of miR-140-5p in vascular aging (created originally by authors through biorender.com).
miR-140-5p and arterial stiffness
Arterial stiffness represents a composite measure of age-associated changes in vascular structure and function,
integrating endothelial dysfunction with medial layer remodeling of the arterial
wall [3, 39, 40].
Central to the development of arterial stiffness is the VSMC phenotypic transition from a quiescent,
contractile state toward a synthetic, proliferative, and sometimes calcifying phenotype, accompanied by ECM imbalance—particularly increased collagen deposition, elastin fragmentation, and fibrosis. Although most research to date has not focused exclusively on miR-140-5p in human vascular aging, several experimental studies provide mechanistic insights into how miR-140-5p influences VSMC behavior and ECM dynamics relevant to arterial stiffness. miR-140-5p has been shown to regulate VSMC viability, migration, and apoptosis in response to oxidized lipoprotein challenge, a key atherogenic stimulus that overlaps with mechanisms of vascular remodeling. In human VSMCs treated with ox-LDL, increased miR-140-5p expression was associated with enhanced proliferation and migration and reduced apoptosis, at least partly through direct targeting of roundabout guidance receptor 4 (ROBO4) gene (Figure 1) [19].
In contrast, other evidence indicates that miR-140-5p can attenuate maladaptive VSMC transitions implicated in stiffness and calcification. In vitro studies demonstrate that miR-140-5p
suppresses VSMC osteogenic differentiation by targeting regulators such as SATB2 [30].
Overexpression of miR-140-5p in this context reduces markers of calcification and ECM mineralization,
while its downregulation promotes these transitions [30].
Although the dual roles of miR-140-5p in both proliferative responses and calcification highlight context-dependent effects, these pathways converge
on key structural determinants of arterial mechanics, namely ECM composition and VSMC phenotype stability.
miRNAs more broadly have been implicated in arterial stiffness via modulation of collagen abundance,
elastin integrity, and VSMC phenotypic plasticity, underscoring the functional plausibility of
miR-140-5p’s involvement in these processes [41]. The
summary of the mechanisms, pathways, and roles of miR-140-5p in vascular aging is presented in Figure 1[19].
miR-140-5p and PWV
PWV integrates endothelial function and arterial wall structure and is the clinical gold-standard measure of large-artery stiffness [5]. Increased PWV reflects cumulative alterations in NO-dependent vasorelaxation, ECM composition (collagen–elastin balance), VSMC phenotype, and medial calcification [42]. miR-140-5p can influence PWV by modulating endothelial homeostasis and NO bioavailability [35]; miR-140-5p affects functional vasorelaxation. miRNA networks are established regulators of eNOS/NO signaling and endothelial oxidative stress, where dysregulated miRNAs correlate with impaired FMD and PWV in human studies [35]. Although strand-specific longitudinal data for miR-140-5p are limited, experimental evidence shows miR-140-5p participates in endothelial stress responses (oxidative and metabolic), which plausibly reduce NO bioavailability when miR-140-5p is dysregulated [35]. In addition, miR-140-5p regulates VSMC phenotype and ECM homeostasis, as suggested by in vitro studies evaluating the effects on target genes such as ROBO4 and SATB2 [19]. Taken together,these functional (endothelial/NO) and structural (VSMC/ECM) mechanisms provide a biologically plausible pathway by which declining miR-140-5p increases PWV, with PWV representing an integrated downstream phenotype of cumulative miR-140-5p–related vascular damage rather than an isolated molecular readout [43].
miR-140-5p in modulating cellular senescence and the senescence-associated secretory phenotype
Cellular senescence is the permanent cessation of proliferation accompanied by
resistance to apoptosis and the acquisition of the senescence-associated secretory
phenotype (SASP). This condition is a major driver of age-related vascular deterioration.
Senescent endothelial cells and VSMCs accumulate in arterial walls with age and contribute
to vascular dysfunction through two mechanisms: loss of regenerative function and secretion
of inflammatory mediators and proteases that damage vascular
structure [25]. Recent evidence has revealed that
miR-140-5p is a negative regulator of senescence whose expression decreases in aging cells
and whose manipulation can modulate the onset and manifestation of senescence.
Senescence is characterized by the activation of the cyclin-dependent kinase inhibitors p16INK4a (CDKN2A) and p21CIP1/WAF1 (CDKN1A),
which induce permanent cell cycle arrest. Studies of IL-1β-induced senescent chondrocytes have
shown that miR-140 overexpression significantly suppresses p16INK4a and p21CIP1/WAF1 expression,
accompanied by reduced senescence-associated β-galactosidase (SA-β-gal) activity and maintenance
of proliferative capacity. Conversely, the inhibition of miR-140 increased the expression of both
senescence markers [44]. Pathway analysis revealed that
miR-140-5p targets components of the Notch (JAG1, NUMBL) and PI3K/AKT (IGF1R, TLR4)
pathways that regulate p16 and p21 expression [10].
Although these data are derived from chondrocytes, their relevance to vascular cells is supported by
the observation that miR-140-5p is expressed in endothelial cells and VSMCs and that its downstream
targets (TLR4 and IGF1R) have been validated in both vascular cell types [10, 45]. However,
further research is needed to confirm the direct regulation of p16/p21 by miR-140-5p in endothelial
cells and VSMCs undergoing senescence.
The SASP encompasses a broad spectrum of proinflammatory cytokines (IL-6, IL-8, and TNF-α),
chemokines (MCP-1), and proteases (matrix metalloproteinases, MMPs) secreted by senescent
cells. These factors create a microenvironment that promotes chronic inflammation
(inflammaging), ECM degradation, and dysfunction of surrounding vascular cells. miR-140-5p
suppresses SASP secretion by targeting key components of the NF-κB activation pathway.
Tumor necrosis factor receptor-associated factor 6 (TRAF6), an adaptor protein that activates
the NF-κB and AP-1 pathways, is a direct target of miR-140-5p, as validated by a luciferase
reporter assay. The overexpression of miR-140 suppresses TRAF6 expression and
reduces the secretion of IL-6, TNF-α, IL-1β, and IFN-γ in infected macrophages [46].
Toll-like receptor 4 (TLR4), a molecular pattern recognition receptor that activates NF-κB,
is also a validated target of miR-140-5p [45, 47]. Although further validation in
vascular-specific contexts is still needed.
In synovial fibroblasts, overexpression of miR-140-5p suppresses TLR4 expression and reduces
IL-6 and IL-8 secretion [45]. In macrophages exposed to
ox-LDL, a miR-140-5p mimic reduces TLR4 expression, decreases oxidative stress, and suppresses
apoptosis [47]. High-mobility group box 1 (HMGB1),
an alarmin that functions as a component of SASP, is another direct target of miR-140-5p. miR-140-5p
suppresses HMGB1 expression and reduces TNF-α, IL-6, MMP-1, and MMP-3 secretion in inflamed
chondrocytes. This effect is mediated through the inhibition of HMGB1 downstream of the
PI3K/AKT pathway [48]. MMP-13 and ADAMTS-5,
proteases that degrade the ECM and contribute to pathological vascular remodeling, are also validated
direct targets of miR-140-5p [49]. These findings indicate
that miR-140-5p suppresses the SASP through multitarget inhibition of the TRAF6/TLR4/NF-κB pathway.
Decreased miR-140-5p levels in aging vascular cells suppress this pathway, amplifying SASP secretion
and contributing to chronic vascular inflammation and the progression of arterial stiffening.
The persistent DNA damage response (DDR) is a strong inducer of senescence. The accumulation of DNA damage
in senescent vascular cells activates histone H2AX phosphorylation (γ-H2AX) and ATM/ATR kinases,
triggering cell cycle arrest and senescent phenotypes. miR-140 suppresses γ-H2AX expression in
senescence-induced chondrocytes, indicating attenuation of DDR signaling [44]. Furthermore,
miR-140 targets flap endonuclease 1 (FEN1), an enzyme involved in DNA repair, and
downregulation of miR-140 increases the accumulation of DNA damage foci in cancer
cells [50]. Whether miR-140-5p modulates the DDR in
endothelial cells and VSMCs requires direct investigation. However, γ-H2AX expression is increased
in the aortas of aged mice and endothelial cells subjected to chronic oxidative stress,
providing a basis for the role of miR-140-5p in vascular maintenance. Therapeutic strategies targeting
miR-140-5p should consider its effects on senescence and the SASP as relevant mechanistic endpoints,
alongside measurements of FMD and PWV as functional
outcomes (Table 2).
Table 2.
Molecular targets of miR-140-5p in senescence and the SASP.
| Molecular targets | Function | Regulatory mechanism | Biological effects | Validation methods | References |
|---|---|---|---|---|---|
| p16INK4a (CDKN2A) | Cell cycle arrest | Suppresses p16 expression | Delaying senescence | qRT-PCR, Western blot | [44] |
| TRAF6 | NF-κB/AP-1 adaptor | Target 3'UTR → ↓ TRAF6 | ↓ IL-6, TNF-α, IL-1β | Luciferase reporter | [46] |
| TLR4 | Inflammatory receptor | Target 3'UTR → ↓ TLR4 | ↓ IL-6, IL-8 | Luciferase reporter | [45] |
| HMGB1 | Alarmin/SASP factor | Target 3'UTR → ↓ HMGB1 | ↓ TNF-α, IL-6, MMP-1, MMP-3 | Luciferase reporter | [48] |
| MMP-13 | ECM protease | Target 3'UTR → ↓ MMP-13 | ↓ matrix degradation | Luciferase reporter | [51] |
| ADAMTS-5 | Aggrecanase | Target 3'UTR → ↓ ADAMTS-5 | ↓ proteoglycan degradation | Luciferase reporter | [49] |
| FEN1 | DNA repair enzyme | Target 3'UTR → ↓ FEN1 | DNA repair disruption | Luciferase reporter | [50] |
Note: SA-β-gal: senescence-associated β-galactosidase, DDR: DNA damage response, IF: immunofluorescence, ECM: extracellular matrix.
Translational and clinical implications
Recent advances in circulating microRNA profiling support the translational relevance of microRNAs as biomarkers of subclinical vascular dysfunction, with multiple studies demonstrating that specific circulating miRNA signatures map to arterial stiffness traits and other early vascular phenotypes, raising the possibility that miR-140-5p may contribute to early detection of vascular aging in selected disease contexts. The integration of mechanistic evidence from epigenetic regulation, cellular senescence, and SASP revealed convergent pathways linking miR-140-5p dysregulation to the clinical parameters of vascular aging. Table 3 summarizes the mechanistic pathways linking miR-140-5p dysregulation at the molecular level with measurable clinical parameters of vascular aging, demonstrating how epigenetic changes and senescence converge to decrease FMD and increase PWV. A direct relationship between miR-140-5p levels and FMD/PWV measurements in human cohorts has not yet been documented. The translational pathways presented in Table 3 were extrapolated from various sources on the basis of (1) experimental evidence of molecular target regulation by miR-140-5p, (2) the well-established role of senescence, SASP, and ECM degradation in vascular pathophysiology, and (3) documented correlations between inflammatory markers, MMPs, and arterial stiffness in clinical studies. Current conclusions are still based on potential mechanistic and indirect evidence, thus needed further studies to conclude robustly.
Table 3.
Integration of miR-140-5p dysregulation with clinical indices of vascular aging.
| Molecular level | Cellular effects | Vascular tissue changes | Clinical phenotype | Measurement parameters | References |
|---|---|---|---|---|---|
| Hypermethylation of the WWP2-miR-140 promoter → ↓ miR-140-5p | Decreased biogenesis of mature miR-140-5p | Loss of proinflammatory target suppression | Endothelial dysfunction | ↓ FMD | [21, 22] |
| ↓ miR-140-5p → ↑ p16INK4a, p21CIP1 | Senescence ECs and VSMCs | Accumulation of senescent cells in the arterial wall | Decreased NO-dependent vasodilation | ↓ FMD | [25, 44] |
| ↓ miR-140-5p → ↑ TRAF6, TLR4, HMGB1 | Activation of NF-κB → SASP | Secretion of IL-6, IL-8, TNF-α (chronic inflammation) | Endothelial dysfunction, arterial stiffening | ↓ FMD, ↑ PWV | [45, 46, 48] |
| ↓ miR-140-5p → ↑ MMP-13, ADAMTS-5 | ECM degradation | Elastin fragmentation, collagen deposition | Decreased arterial compliance | ↑ PWV | [49, 51] |
| ↓ miR-140-5p → ↑ ROBO4, loss of SATB2 suppression | VSMC phenotype switching | Osteoblastic transition, medial calcification | Increased arterial stiffness | ↑ PWV | [19, 30] |
| ceRNA sequestration (H19 ↑) → ↓ miR-140-5p bioavailability | Loss of pro-calcification target suppression | Progressive vascular calcification | Arterial stiffening | ↑ PWV | [30] |
Note: EC: endothelial cell; VSMC: vascular smooth muscle cell; ECM: extracellular matrix; NO: nitric oxide; FMD: flow-mediated dilation; PWV: pulse wave velocity; SASP: senescence-associated secretory phenotype.
Although miR-140-5p is generally described as a regulator of vascular homeostasis,
which indicates that its biological effects are often highly context-dependent and
potentially bidirectional among conditions. Under physiological or moderate stress
conditions, it appears to produce protective roles through modulating inflammatory
signaling, limits ECM degradation, and also attenuating cellular senescence pathways.
However, among pathological conditions, where sustained oxidative stress or metabolic
disturbance occur, miR-140-5p exacerbate redox imbalance, contributing to vascular
dysfunction through interactions with the Nrf2/SIRT2 axis. Thus, it should be interpreted
according to context, which implies that therapeutic strategies targeting miR-140-5p will
require careful consideration of tissue specificity, timing, and dosage.
With recent progress in miRNA-based therapeutics, this early evidence suggests that
targeted modulation of miR-140-5p using mimics or antagomirs represents a feasible
experimental strategy to test whether restoring miR-140-5p can preserve endothelial
function and limit arterial stiffening. However, available studies also highlight
context-dependent effects, with miR-140-5p promoting oxidative stress and hypertension
in certain models, underscoring the need for strand-specific, tissue-targeted approaches
and careful dose optimization. To advance clinical translation, priority should be given
to cross-sectional studies integrating plasma or tissue miR-140-5p measurements with FMD
and carotid-femoral PWV in age-stratified cohorts, longitudinal studies assessing whether
baseline miR-140-5p predicts PWV progression and cardiovascular events, and preclinical
interventional studies in which vascular compliance or PWV are prespecified endpoints.
Limitations and research gaps
Despite the mechanistic plausibility, direct human evidence linking miR-140-5p to clinically measured vascular aging indices, particularly flow-mediated dilation and PWV, especially through cohort studies, remains limited. Thus, the proposed relationships among them were biologically plausible which has not been clinically validated and should therefore be cautiously interpreted. Most available data are derived from in vitro experiments or disease-oriented animal models, which may not fully capture the gradual and multifactorial nature of physiological vascular aging. In addition, existing studies are largely cross-sectional and do not allow assessment of temporal relationships between miR-140-5p expression and progression of vascular dysfunction. Furthermore, the majority of mechanistic evidence concerning the regulation of p16/p21 and SA-β-gal by miR-140-5p originates from chondrocyte models. While the same downstream targets (TLR4 and TRAF6) have been validated in vascular cells, further research is required to confirm the effects of miR-140-5p on endothelial cell and VSMC senescence. Strand-specific effects of miR-140-5p further complicate interpretation, as context-dependent and sometimes opposing biological actions have been reported across different cellular environments and pathological states. Importantly, it is not yet established whether modulation of miR-140-5p can causally improve endothelial function, reduce arterial stiffness, or alter PWV in vivo. Addressing these gaps will require well-phenotyped, age-stratified human cohorts, longitudinal designs integrating molecular and vascular functional data, and carefully controlled interventional studies that account for tissue specificity, dosage, and off-target effects of miR-140-5p modulation.
Conclusions
Emerging evidence, mostly from animal studies, indicates that miR-140-5p influences endothelial NO bioavailability, oxidative and inflammatory signaling, VSMC phenotype, and ECM homeostasis. Through these integrated effects, age-related dysregulation of miR-140-5p provides a plausible molecular link between cellular stress responses and clinically measurable vascular aging phenotypes. The integration of epigenetic, senescence, and SASP evidence revealed that the decrease in miR-140-5p during vascular aging is not a passive phenomenon but rather the result of coordinated changes in gene regulation, microRNA biogenesis, and noncoding RNA networks. At the same time, current evidence underscores important limitations, particularly the scarcity of longitudinal human data directly connecting miR-140-5p levels with FMD and PWV. Future studies combining well-phenotyped aging cohorts, longitudinal vascular assessments, and strand-specific interventional approaches in well-designed human studies will be essential to clarify whether miR-140-5p functions primarily as a biomarker, a mechanistic driver, or both.
Declarations
Availability of data and materials
Not applicable.
Financial support and sponsorship
None.
Conflicts of interest
Not applicable.
Ethical approval and informed consent
Not applicable.
Consent for publication
Not applicable.
AI and AI-assisted tools statement
No AI tools were used in the preparation of this article.
References
1. Zhang S, Xia B, Kalionis B, Li H, Zhang X, Zhang X, et al. The role and mechanism of vascular aging in geriatric vascular diseases. Aging Dis, 2024, 16(4): 2237-2249. [Crossref]
2. González L, Romero-Orjuela S, Rabeya F, del Castillo V, & Echeverri D. Age and vascular aging: an unexplored frontier. Front Cardiovasc Med, 2023, 10: 1278795. [Crossref]
3. Oliveira A, Cunha P, Vitorino P, Souza A, Deus G, Feitosa A, et al. Vascular aging and arterial stiffness. Arq Bras Cardiol, 2022, 119(4): 604-615. [Crossref]
4. Heiss C, Rodriguez-Mateos A, Bapir M, Skene S, Sies H, & Kelm M. Flow-mediated dilation reference values for evaluation of endothelial function and cardiovascular health. Cardiovasc Res, 2023, 119(1): 283-293. [Crossref]
5. Heffernan K, Stoner L, London A, Augustine J, & Lefferts W. Estimated pulse wave velocity as a measure of vascular aging. PLoS One, 2023, 18(1): e0280896. [Crossref]
6. Lin Z, Ding Q, Li X, Feng Y, He H, Huang C, et al. Targeting epigenetic mechanisms in vascular aging. Front Cardiovasc Med, 2022, 8: 806988. [Crossref]
7. Ma S, Xie X, Yuan R, Xin Q, Miao Y, Leng S, et al. Vascular aging and atherosclerosis: a perspective on aging. Aging Dis, 2025, 16(1): 33-48. [Crossref]
8. Wang G, Luo Y, Gao X, Liang Y, Yang F, Wu J, et al. MicroRNA regulation of phenotypic transformations in vascular smooth muscle: relevance to vascular remodeling. Cell Mol Life Sci, 2023, 80(6): 144-156. [Crossref]
9. Sieland J, Niederer D, Engeroff T, Vogt L, Troidl C, Schmitz-Rixen T, et al. Changes in miRNA expression in patients with peripheral arterial vascular disease during moderate- and vigorous-intensity physical activity. Eur J Appl Physiol, 2023, 123(3): 645-654. [Crossref]
10. Toury L, Frankel D, Airault C, Magdinier F, Roll P, & Kaspi E. miR-140-5p and miR-140-3p: key actors in aging-related diseases? Int J Mol Sci, 2022, 23(19): 11439. [Crossref]
11. Li J, Jin Y, Cao Y, & Wu G. MiR-140-5p exerts a protective function in pregnancy-induced hypertension via mediating TGF-β/Smad signaling pathway. Hypertens Pregnancy, 2022, 41(2): 116-125. [Crossref]
12. Qiao P, Zhu J, Lu X, Jin Y, Wang Y, Shan Q, et al. miR-140-3p suppresses the proliferation and migration of macrophages. Genet Mol Biol, 2022, 45(2): e20210160. [Crossref]
13. Turko R, Hajja A, Magableh A, Omer M, Shafqat A, Khan M, et al. The emerging role of miRNAs in biological aging and age-related diseases. Non-coding RNA Res, 2025, 13: 131-152. [Crossref]
14. Machaj F, Chmielewska-Jeznach M, Koryszewska-Bagińska A, Malinowski D, Pawlik A, & Olędzka G. MicroRNAs as biomarkers and therapeutic targets in rheumatoid arthritis. Int J Mol Sci, 2025, 26(20): 9950. [Crossref]
15. Núñez-Carro C, Blanco-Blanco M, Villagrán-Andrade K, Blanco F, & de Andrés M. Epigenetics as a therapeutic target in osteoarthritis. Pharmaceuticals, 2023, 16(2): 156-176. [Crossref]
16. Fu C, Peng J, Ling Y, Zhao H, Zhao Y, Zhang X, et al. Apigenin inhibits angiogenesis in retinal microvascular endothelial cells through regulating of the miR-140-5p/HDAC3-mediated PTEN/PI3K/AKT pathway. BMC Ophthalmol, 2023, 23(1): 302-316. [Crossref]
17. Shang Z, & Huang K. Cyclic stretch and oxidative stress induced miRNAs in vascular remodeling. Biophys Rep, 2025, 11: 1-9. [Crossref]
18. Toury L, Frankel D, Nael S, Abaji M, Le Goff L, Basset M, et al. miR-140-5p overexpression contributes to oxidative stress and mitochondrial dysfunction in Hutchinson-Gilford Progeria syndrome fibroblasts through Nrf2 pathway. Aging Cell, 2025, 24(12): e70276. [Crossref]
19. Luo Y, Li Y, Peng H, & Zhao Y. miR-140-5p regulates vascular smooth muscle cell viability, migration and apoptosis by targeting ROBO4 gene expression in atherosclerosis. Mol Med Rep, 2021, 23(3): 213-226. [Crossref]
20. Yang J, Qin S, Yi C, Ma G, Zhu H, Zhou W, et al. MiR-140 is co-expressed with Wwp2-C transcript and activated by Sox9 to target Sp1 in maintaining the chondrocyte proliferation. FEBS Lett, 2011, 585(19): 2992-2997. [Crossref]
21. Roberts J, Boldvig O, Aubourg G, Kanchanapally S, Dehan D, Rice S, et al. Specific isoforms of the ubiquitin ligase gene WWP2 are targets of osteoarthritis genetic risk via a differentially methylated DNA sequence. Arthritis Res Ther, 2024, 26(1): 78-88. [Crossref]
22. Papathanasiou I, Trachana V, Mourmoura E, & Tsezou A. DNA methylation regulates miR-140-5p and miR-146a expression in osteoarthritis. Life Sci, 2019, 228: 274-284. [Crossref]
23. Xu H, Li S, & Liu Y. Roles and mechanisms of DNA methylation in vascular aging and related diseases. Front Cell Dev Biol, 2021, 9: 699374. [Crossref]
24. Duru N, Gernapudi R, Zhang Y, Yao Y, Lo P, Wolfson B, et al. Nrf2/miR-140 signaling confers radioprotection to human lung fibroblasts. Cancer Lett, 2015, 369(1): 184-191. [Crossref]
25. Colpani O, & Spinetti G. MicroRNAs orchestrating senescence of endothelial and vascular smooth muscle cells. Vasc Biol, 2019, 1(1): H75-H81. [Crossref]
26. Liu Q, Ren K, Liu S, Li W, Huang C, & Yang X. MicroRNA-140-5p aggravates hypertension and oxidative stress of atherosclerosis via targeting Nrf2 and Sirt2. Int J Mol Med, 2019, 43(2): 839-849. [Crossref]
27. Ungvari Z, Tucsek Z, Sosnowska D, Toth P, Gautam T, Podlutsky A, et al. Aging-induced dysregulation of Dicer1-dependent microRNA expression impairs angiogenic capacity of rat cerebromicrovascular endothelial cells. J Gerontol A Biol Sci Med Sci, 2013, 68(8): 877-891. [Crossref]
28. Hartmann P, Zhou Z, Natarelli L, Wei Y, Nazari-Jahantigh M, Zhu M, et al. Endothelial Dicer promotes atherosclerosis and vascular inflammation by miRNA-103-mediated suppression of KLF4. Nat Commun, 2016, 7: 10521. [Crossref]
29. Braun H, Hauke M, Ripperger A, Ihling C, Fuszard M, Eckenthaler R, et al. Impact of DICER1 and DROSHA on the angiogenic capacity of human endothelial cells. Int J Mol Sci, 2021, 22(18): 9855-9869. [Crossref]
30. Xu F, Zhong J, Guo B, Lin X, Wu F, Li F, et al. H19 promotes osteoblastic transition by acting as ceRNA of miR-140-5p in vascular smooth muscle cells. Front Cell Dev Biol, 2022, 10: 774363. [Crossref]
31. Zhao Z, Zhang M, Liao L, Zou J, Wang G, Wan X, et al. Long non-coding RNA PCA3 inhibits lipid accumulation and atherosclerosis through the miR-140-5p/RFX7/ABCA1 axis. Biochim Biophys Acta Mol Cell Biol Lipids, 2021, 1866(5): 158904. [Crossref]
32. Fan P, Chen Z, Tian P, Liu W, Jiao Y, Xue Y, et al. miRNA biogenesis enzyme Drosha is required for vascular smooth muscle cell survival. PLoS One, 2013, 8(4): e60888. [Crossref]
33. Wang X, & He B. Endothelial dysfunction: molecular mechanisms and clinical implications. MedComm, 2024, 5(8): e651. [Crossref]
34. Hu Y, Chen M, Wang M, & Li X. Flow-mediated vasodilation through mechanosensitive G protein-coupled receptors in endothelial cells. Trends Cardiovasc Med, 2022, 32(2): 61-70. [Crossref]
35. Nemecz L, Alexandru N, Tanko G, & Georgescu A. Role of microRNA in endothelial dysfunction and hypertension. Curr Hypertens Rep, 2016, 18(12): 87-96. [Crossref]
36. Zhang Q, Long J, Li N, Ma X, & Zheng L. Circ_CLASP2 regulates high glucose-induced dysfunction of human endothelial cells through targeting miR-140-5p/FBXW7 axis. Front Pharmacol, 2021, 12: 594793. [Crossref]
37. Minjares M, Wu W, & Wang J. Oxidative stress and microRNAs in endothelial cells under metabolic disorders. Cells, 2023, 12(9): 1341-1352. [Crossref]
38. Zhang B, Li Q, Lu Y, Wang W, Tian M, Guo J, et al. KCNQ1OT1/miR-140-5p/PTP4A3 axis is involved in endosulfan-induced vascular endothelial cell migration linking to atherosclerosis. Toxicol Lett, 2025, 412: 44-54. [Crossref]
39. Lee G, & Kim H. Incremental value of the measures of arterial stiffness in cardiovascular risk assessment. Rev Cardiovasc Med, 2022, 23(1): 6-17. [Crossref]
40. Xuereb RA, Magri C, & Xuereb RG. Arterial stiffness and its impact on cardiovascular health. Curr Cardiol Rep, 2023, 25(10): 1337-1349. [Crossref]
41. Nanoudis S, Pikilidou M, Yavropoulou M, & Zebekakis P. The role of microRNAs in arterial stiffness and arterial calcification. An update and review of the literature. Front Genet, 2017, 8: 209-215. [Crossref]
42. Marshall A, Neikirik K, Afolabi J, Mwesigwa N, Shao B, Kirabo A, et al. Update on the use of pulse wave velocity to measure age-related vascular changes. Curr Hypertens Rep, 2024, 26(3): 131-140. [Crossref]
43. Nanoudis S, Yavropoulou M, Tsachouridou O, Pikilidou M, Pilalas D, Kotsa K, et al. Circulating microRNAs related to arterial stiffness in adults with HIV infection. Viruses, 2024, 16(12): 1945-1957. [Crossref]
44. Si H, Yang T, Li L, Tian M, Zhou L, Li D, et al. miR-140 attenuates the progression of early-stage osteoarthritis by retarding chondrocyte senescence. Mol Ther Nucleic Acids, 2020, 19: 15-30. [Crossref]
45. Li H, Guan S, Lu Y, & Wang F. MiR-140-5p inhibits synovial fibroblasts proliferation and inflammatory cytokines secretion through targeting TLR4. Biomed Pharmacother, 2017, 96: 208-214. [Crossref]
46. Li X, Huang S, Yu T, Liang G, Liu H, Pu D, et al. MiR-140 modulates the inflammatory responses of Mycobacterium tuberculosis-infected macrophages by targeting TRAF6. J Cell Mol Med, 2019, 23(8): 5642-5653. [Crossref]
47. Liu H, Mao Z, Zhu J, Shen M, & Chen F. MiR-140-5p inhibits oxidized low-density lipoprotein-induced oxidative stress and cell apoptosis via targeting toll-like receptor 4. Gene Ther, 2021, 28(7): 413-421. [Crossref]
48. Wang Y, Shen S, Li Z, Li W, & Wang X. MiR-140-5p affects chondrocyte proliferation, apoptosis, and inflammation by targeting HMGB1 in osteoarthritis. Inflamm Res, 2020, 69(1): 63-73. [Crossref]
49. Miyaki S, Nakasa T, Otsuki S, Grogan S, Higashiyama R, Inoue A, et al. MicroRNA-140 is expressed in differentiated human articular chondrocytes and modulates inter leukin-1 responses. Arthritis Rheum, 2009, 60(9): 2723-2730. [Crossref]
50. Lu X, Liu R, Wang M, Kumar A, Pan F, He L, et al. MicroRNA-140 impedes DNA repair by targeting FEN1 and enhances chemotherapeutic response in breast cancer. Oncogene, 2020, 39(1): 234-247. [Crossref]
51. Tardif G, Hum D, Pelletier J, Duval N, & Martel-Pelletier J. Regulation of the IGFBP-5 and MMP-13 genes by the microRNAs miR-140 and miR-27a in human osteoarthritic chondrocytes. BMC Musculoskelet Disord, 2009, 10(1): 148. [Crossref]