Open Access | Review
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
mTOR signaling in aging and longevity: therapeutic opportunities
* Corresponding author: Gaurav N. Kasar
Department of Pharmacology, VADP’s VPD Institute of Pharmacy, Shirsondi, (Malegaon), Department of Pharmacology, SNJB’s
SSDJ College of Pharmacy, Chandwad (Nashik), India.
Email: gauravkasar008@gmail.com
Received: 30 April 2026 / Revised: 21 May 2026 / Accepted: 11 June 2026 / Published: 30 June 2026
DOI: 10.31491/APT.2026.06.216
Abstract
Aging is a complex biological process characterized by the gradual decline of physiological functions and an increased susceptibility to chronic diseases, including cancer, cardiovascular disorders, metabolic syndromes, and neurodegenerative conditions. Among the major molecular pathways associated with aging, the mechanistic target of rapamycin (mTOR) signaling pathway has emerged as a central regulator of cellular growth, metabolism, and longevity. mTOR integrates signals from nutrients, growth factors, and cellular energy status to regulate essential cellular processes such as protein synthesis, autophagy, stress response, and metabolic homeostasis.
This review explores the role of mTOR signaling in aging and age-related diseases by summarizing findings from recent experimental and clinical studies. Evidence suggests that dysregulated mTOR activity contributes to several hallmarks of aging, including cellular senescence, impaired autophagy, metabolic imbalance, mitochondrial dysfunction, and stem cell exhaustion. Persistent activation of mTOR has been strongly associated with the progression of age-related disorders, whereas controlled inhibition of mTOR signaling has been shown to extend lifespan and improve healthspan in various model organisms.
Several therapeutic and lifestyle-based interventions targeting mTOR have demonstrated promising anti-aging effects. Pharmacological agents such as rapamycin and its analogues regulate mTOR activity and delay the onset of aging-associated diseases. In addition, non-pharmacological strategies including calorie restriction, intermittent fasting, regular physical exercise, and natural bioactive compounds have shown potential in modulating mTOR signaling and improving metabolic health.
Recent advances in geroscience further highlight emerging approaches such as combination therapies targeting multiple longevity pathways, integration of senolytic strategies, and precision medicine-based interventions designed to personalize anti-aging treatments. Although preclinical findings are encouraging, further long-term clinical studies are required to evaluate the efficacy, safety, and translational potential of mTOR-targeted therapies. A deeper understanding of mTOR signaling in aging biology may contribute to the development of innovative therapeutic strategies for promoting healthy aging and preventing age-related diseases.
Keywords
Aging, mTOR signaling, longevity, autophagy, anti-aging therapeutics
Introduction
The world’s population is aging quickly, which presents serious problems for socioeconomic institutions and healthcare systems everywhere. The World Health Organization projects that by 2050, there will be more than 2 billion people over the age of 60 [1]. Numerous chronic illnesses, such as cancer, metabolic syndromes, neurological problems, and cardiovascular diseases, are primarily associated with aging. Global morbidity and mortality are significantly impacted by these illnesses’ rising prevalence. Crucially, aging is now understood to be a complicated biological process marked by growing molecular and cellular degradation rather than just a temporal event [2]. Damage accumulation eventually causes tissues and organs to deteriorate functionally, making them more susceptible to illness. In order to increase longevity and postpone the development of age-related illnesses, biomedical research has turned its attention to comprehending the molecular mechanisms behind aging [3].
Researchers have created the framework known as the hallmarks of aging, which describes the basic biological processes that lead to aging, in order to better understand the molecular foundation of aging [2]. Genomic instability, telomere attrition, epigenetic changes, loss of proteostasis, mitochondrial failure, cellular senescence, stem cell exhaustion, and altered intercellular communication are among the interrelated pathways identified by this conceptual paradigm. In order to highlight the dynamic and complex nature of aging biology, more recent revisions of this framework have broadened the notion to incorporate new elements including dysregulated nutrient sensing and chronic inflammation [4]. These characteristics interact in intricate networks that gradually compromise physiological function and cellular equilibrium rather than
functioning independently. Understanding these processes has provided important insights into potential therapeutic targets that may slow aging and prevent multiple age-related pathologies simultaneously [5].
Nutrient-sensing pathways are one of the biological processes that affect aging, and they are especially important for controlling lifespan and metabolic balance. Cells are able to adjust to environmental changes in food availability and energy status due to these evolutionarily conserved signaling mechanisms. AMP-activated protein kinase (AMPK), sirtuins, insulin-like growth factor 1 (IGF-1) signaling, and the mechanistic target of rapamycin (mTOR) pathway are important nutrient-sensing systems. In reaction to nutritional levels, these pathways regulate cellular functions, including energy metabolism, autophagy, stress tolerance, and protein synthesis. Modulation of nutrient-sensing pathways can dramatically increase lifespan and improve metabolic health, according to experimental data from a variety of model animals, including yeast, nematodes, fruit flies, and mammals [3]. Interventions such as caloric restriction and intermittent fasting exert many of their beneficial effects by influencing these molecular networks, highlighting their importance in the regulation of aging and longevity [6, 7].
The mTOR signaling pathway has become a key integrator of cellular growth, metabolism, and aging processes among these nutrient-sensing regulators. Protein synthesis, lipid metabolism, cytoskeletal architecture, and autophagy are just a few of the cellular processes that mTOR controls through two different multiprotein complexes, mechanistic target of rapamycin complex 1 (mTORC1) and mechanistic target of rapamycin complex 2 (mTORC2) [6]. Accelerated aging and a number of age-related illnesses, including neurodegeneration, cancer, and metabolic disorders, have been connected to dysregulation or persistent activation of mTOR signaling [7]. On the other hand, in a number of experimental models, pharmacological inhibition of mTOR with drugs like rapamycin has been demonstrated to increase longevity and improve healthspan. These discoveries have made mTOR one of the most promising biological targets for therapeutic strategies meant to slow down aging and stop age-related illnesses. Therefore, understanding the mechanisms and regulatory networks of mTOR signaling is essential for the development of novel strategies to promote healthy aging and longevity [8]. In this review, we aim to summarize the molecular mechanisms of mTOR signaling, its role in aging and age-related diseases, and current therapeutic strategies targeting the mTOR pathway to promote healthy aging and longevity.
Structure and components of the mTOR pathway
The highly conserved serine/threonine kinase signaling network known as the mTOR pathway is essential for controlling cellular growth, metabolism, survival, and aging. To coordinate anabolic and catabolic processes, mTOR integrates a variety of intracellular and extracellular cues, such as nutrition availability, growth hormones, energy status, and cellular stress [9]. From a structural perspective, mTOR serves as the catalytic core of two different multiprotein complexes called mTORC1 and mTORC2. These complexes have different physiological activities, downstream substrates, regulatory mechanisms, and protein compositions. mTORC2 is principally engaged in the regulation of cell survival, cytoskeletal architecture, and metabolic signaling, whereas mTORC1 predominantly controls protein synthesis, autophagy, and metabolic homeostasis in response to nutrient signals. Understanding these complexes’ structural organization and biological functions is crucial since dysregulation of these complexes has been linked to a number of clinical illnesses, including cancer, neurological disorders, metabolic diseases, and aging-related dysfunctions [10, 11]. The structural organization and major regulatory components of the mTOR signaling pathway are illustrated in Figure 1.
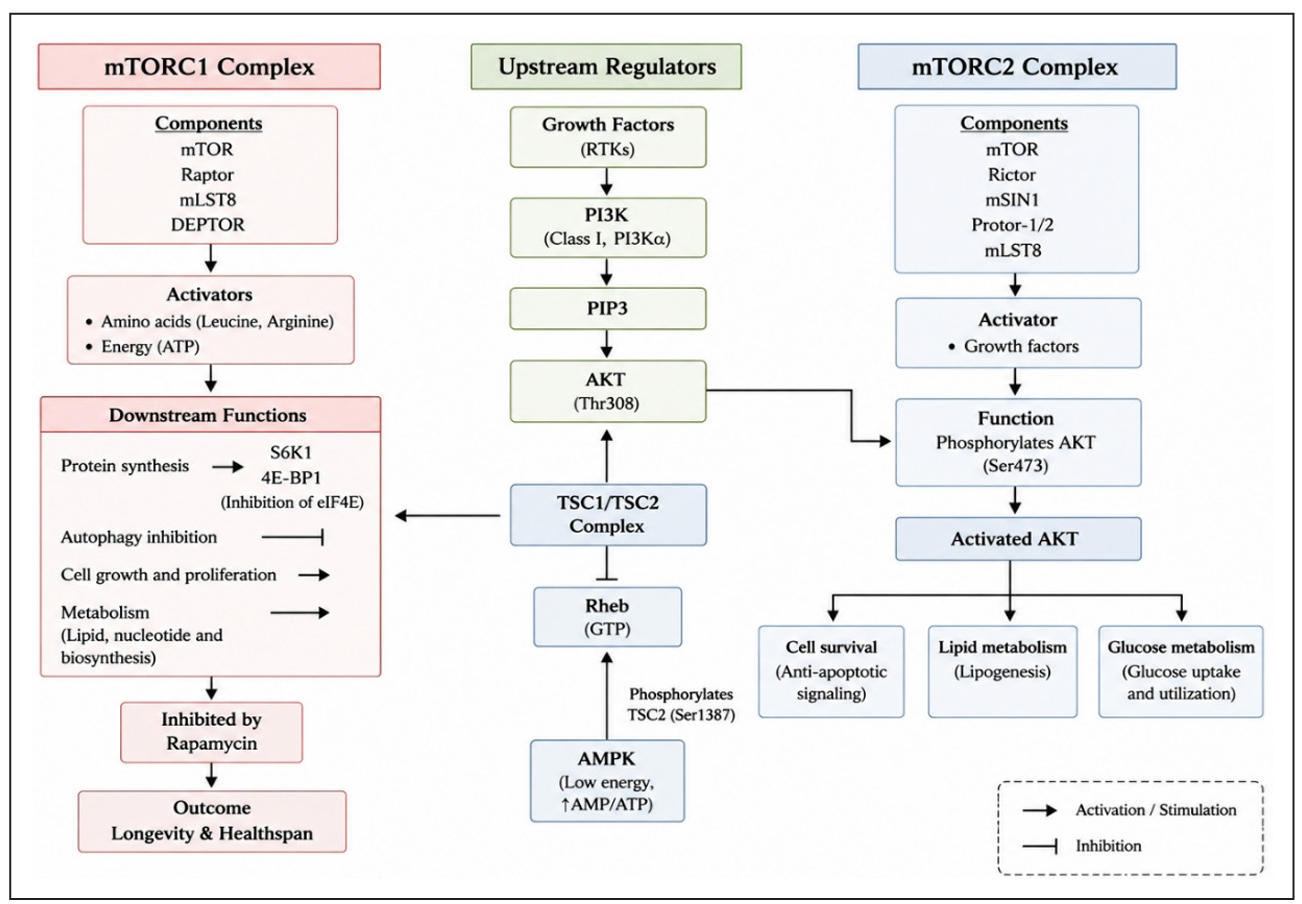
Figure 1. Schematic representation of the mTOR signaling pathway illustrating the structural organization, upstream regulators, and downstream cellular functions of mTORC1 and mTORC2 complexes. mTORC1, composed of mTOR, Raptor, mLST8, and DEPTOR, is primarily activated by amino acids and cellular energy availability and regulates protein synthesis, autophagy inhibition, cell growth, and metabolism through downstream effectors such as S6K1 and 4E-BP1. In contrast, mTORC2, consisting of mTOR, Rictor, mSIN1, Protor-1/2, and mLST8, is mainly activated by growth factors and promotes AKT phosphorylation at Ser473, thereby regulating cell survival, lipid metabolism, and glucose metabolism. The figure further highlights the involvement of the PI3K/AKT and AMPK signaling pathways in modulating mTOR activity through regulation of the TSC1/TSC2 complex and Rheb signaling. Abbreviations: mTOR, mechanistic target of rapamycin; mTORC1, mechanistic target of rapamycin complex 1; mTORC2, mechanistic target of rapamycin complex 2; PI3K, phosphatidylinositol-3-kinase; PIP3, phosphatidylinositol3,4,5-trisphosphate; AKT, protein kinase B; AMPK, AMP-activated protein kinase; TSC1/TSC2, tuberous sclerosis complex 1/2; Rheb, Ras homolog enriched in brain; S6K1, ribosomal protein S6 kinase beta-1; 4E-BP1, eukaryotic translation initiation factor 4E-binding protein 1; RTKs, receptor tyrosine kinases; ATP, adenosine triphosphate; AMP, adenosine monophosphate; mLST8, mammalian lethal with SEC13 protein 8; DEPTOR, DEP domain-containing mTOR-interacting protein; Raptor, regulatory-associated protein of mTOR; Rictor, rapamycin-insensitive companion of mTOR; mSIN1, mammalian stress-activated protein kinase interacting protein 1; Protor-1/2, protein observed with Rictor-1/2.
mTOR complexes
mTORC1
As a key regulator of cell growth and metabolism, the mTORC1 complex is the most researched branch of the mTOR signaling pathway. A number of essential elements make up the complex, including mTOR, proline-rich AKT substrate 40 kDa (PRAS40), DEP domain-containing mTOR-interacting protein (DEPTOR), mammalian lethal with SEC13 protein 8 (mLST8), and regulatory-associated protein of mTOR (Raptor). Among these, Raptor functions as a scaffolding protein that makes it easier to recognize and attract substrates, allowing mTORC1 to phosphorylate important downstream targets involved in cellular growth and protein translation [10]. Nutrients such as amino acids (particularly leucine and arginine), growth factors, and energy availability are the main triggers for mTORC1 activation [11]. When mTORC1 is activated, it phosphorylates downstream effectors that drive mRNA translation and protein synthesis, such as ribosomal protein S6 kinase beta-1 (S6K1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1). Furthermore, autophagy, a cellular degradation process necessary for preserving cellular homeostasis and eliminating damaged organelles, is adversely regulated by mTORC1 [12]. While pharmacological suppression of this complex with drugs like rapamycin has been demonstrated to increase lifespan in numerous experimental organisms, chronic activation of the mTORC1 has been associated with accelerated aging and several age-related illnesses [13].
mTORC2
The second functional arm of the mTOR signaling pathway is represented by the mTORC2 complex, which differs from the mTORC1 in both structure and regulation. mTOR, rapamycin-insensitive companion of mTOR (Rictor), mammalian stress-activated protein kinase interacting protein 1 (mSIN1), protein observed with Rictor-1/2 (Protor-1/2), mLST8, and DEPTOR are the main constituents of mTORC2. In contrast to mTORC1, which predominantly reacts to the availability of nutrients, mTORC2 is largely activated by growth factor signaling pathways and is essential for controlling metabolism, cytoskeletal architecture, and cell survival [11, 12]. The serine/threonine kinase protein kinase B (AKT), which is phosphorylated at Ser473 for complete activation, is one of the most significant downstream substrates of the mTORC2. mTORC2 affects cell survival pathways, lipid synthesis, and glucose metabolism via AKT and other signaling molecules [13]. Long-term exposure to rapamycin may indirectly affect mTORC2 assembly in some cell types, despite the fact that mTORC2 is typically thought to be less vulnerable to acute suppression by rapamycin than mTORC1. Although its function in longevity is still less clear than that of mTORC1, new data indicate that mTORC2 also has a role in aging-related metabolic regulation [14].
Upstream regulators of the mTOR pathway
PI3K/AKT signaling pathway
The PI3K/AKT signaling pathway is one of the key upstream regulators of the mTOR pathway. This pathway is often triggered when external growth stimuli bind to and induce the phosphorylation of cell surface receptor tyrosine kinases (RTKs). Phosphorylated RTKs subsequently recruit and activate class I phosphatidylinositol-3-kinase (class I PI3K), particularly the PI3Kα isoform, which plays a major role in growth factor-mediated signaling. This activation stimulates phosphatidylinositol-3-kinase (PI3K), which catalyzes the conversion of phosphatidylinositol-4,5-bisphosphate (PIP2) into phosphatidylinositol-3,4,5-trisphosphate (PIP3) [15].
AKT, a serine/threonine kinase, is recruited to the plasma membrane following the accumulation of PIP3, where it undergoes phosphorylation and complete activation by phosphoinositide-dependent kinase-1 (PDK1) and mTORC2. Activated AKT subsequently phosphorylates several downstream targets, including the tuberous sclerosis complex (TSC1/TSC2), thereby relieving its inhibitory effect on the mTORC1. As a result, mTORC1 becomes activated and promotes anabolic cellular processes such as protein synthesis, cell growth, and metabolism. This signaling cascade is essential for regulating cellular proliferation and survival and is frequently altered under abnormal conditions such as cancer and metabolic disorders [16].
AMPK
AMPK, a cellular energy sensor, is another important regulator of mTOR signaling. When cellular energy levels are low, as indicated by an elevated AMP/ATP ratio, AMPK becomes activated. To maintain cellular energy homeostasis and metabolic balance, activated AMPK inhibits mTORC1 activity [16]. Protein synthesis and other anabolic processes are suppressed through phosphorylation of the TSC2 complex, particularly at Ser1387, as well as direct phosphorylation of the mTORC1 component Raptor. Through this mechanism, AMPK suppresses excessive energy consumption while promoting catabolic pathways such as fatty acid oxidation and autophagy.
In contrast, under nutrient-rich and growth factor-stimulated conditions, AKT phosphorylates TSC2 at different regulatory sites, leading to inhibition of the TSC1/TSC2 complex and subsequent activation of the mTORC1. Thus, AMPK and AKT exert reciprocal regulatory control over the TSC2 complex under energy-deficient and energy-sufficient conditions, respectively, thereby coordinating the balance between catabolic and anabolic cellular processes. This reciprocal regulation ensures that cell growth, proliferation, and biosynthetic activities occur only when sufficient energy and nutrient availability are present [17].
Growth factors and nutrient availability
The availability of nutrients and growth factors is crucial upstream cues that control mTOR activity. Growth hormones such as insulin and insulin-like growth factor-1 activate RTKs, which subsequently stimulate the PI3K/AKT pathway and promote the mTORC1 activation [15].
Apart from growth factor signaling, amino acids, particularly leucine, arginine, and glutamine, are essential activators of the mTORC1. Specialized intracellular sensors detect amino acid availability and regulate the localization of mTORC1 to the lysosomal surface, where it is activated by the small GTPase Rheb. Glutamine additionally contributes to the mTORC1 regulation through multiple mechanisms, including amino acid exchange via transporters such as SLC1A5 and SLC7A5/SLC3A2, which facilitate intracellular leucine accumulation and subsequent mTORC1 activation [16]. Furthermore, glutaminolysis-derived metabolites support cellular bioenergetics and promote cell cycle progression, thereby linking glutamine metabolism with the mTORC1 signaling [17]. Through these coordinated mechanisms, cells synchronize nutrient availability with anabolic metabolism, protein synthesis, and cellular proliferation. Conversely, nutrient deprivation suppresses mTOR signaling and induces autophagy to recycle intracellular components and maintain cellular homeostasis [18].
Downstream targets of mTOR
S6K1
S6K1 is one of the main downstream targets of the mTORC1. S6K1 is activated when mTORC1 phosphorylates it at particular sites, such as Thr389. A number of proteins involved in the control of mRNA translation, such as ribosomal protein S6, eukaryotic initiation factor 4B (eIF4B), and other translational regulators, are subsequently phosphorylated by activated S6K1. These phosphorylation processes promote cellular development and protein synthesis by improving ribosome biogenesis and translational efficiency. S6K1 signaling plays a crucial role in coordinating cellular growth responses downstream of mTOR activation, as evidenced by its contribution to the regulation of cell size, cell cycle progression, and metabolic pathways [19].
4E-BP1
4E-BP1 is another important downstream effector of the mTORC1. By attaching to the eukaryotic initiation factor 4E (eIF4E) and blocking the development of the eIF4F translation initiation complex, 4E-BP1 acts as a translational repressor in baseline conditions. When mTORC1 is activated, 4E-BP1 is phosphorylated and dissociates from eIF4E. This release starts protein synthesis by enabling eIF4E to take part in cap-dependent mRNA translation. The mTOR-4E-BP1 axis is essential for controlling cell growth, proliferation, and metabolic activity through this method. Numerous illnesses, such as cancer and metabolic disorders, have been linked to dysregulation of this system [20].
Regulation of protein synthesis and metabolism
S6K1 and 4E-BP1 work together to allow mTORC1 to control metabolism and protein synthesis. While phosphorylation of 4E-BP1 encourages cap-dependent translation of growth-related proteins, activation of S6K1 increases translation of mRNAs involved in ribosome synthesis and metabolic enzymes. When combined, these strategies enable mTOR signaling to restrict catabolic pathways like autophagy while promoting anabolic activities including protein synthesis, lipid biosynthesis, and nucleotide synthesis. Thus, by combining upstream signals from nutrients, growth factors, and energy status with downstream translational machinery, the mTOR pathway functions as a master regulator of cellular metabolism and growth [6].
Role of mTOR signaling in aging biology
The mTOR signaling pathway plays a central role in regulating multiple biological processes associated with aging [21]. Through its two complexes, mTORC1 and mTORC2, mTOR integrates signals from nutrients, growth factors, and cellular energy status to control protein synthesis, metabolism, autophagy, and cellular stress responses [22]. The major mechanisms through which dysregulated the mTORC1 signaling contributes to aging, including cellular senescence, senescence-associated secretory phenotype (SASP), impaired autophagy, mitochondrial dysfunction, altered nutrient sensing, and stem cell exhaustion, are illustrated in Figure 2. Dysregulated or chronic activation of mTOR signaling has been strongly associated with several hallmarks of aging, including cellular senescence, impaired autophagy, metabolic imbalance, mitochondrial dysfunction, and stem cell exhaustion [23]. Conversely, inhibition of mTOR activity, particularly by pharmacological agents such as rapamycin, has been shown to extend lifespan and improve healthspan in several experimental organisms, highlighting mTOR as a critical therapeutic target for healthy aging and longevity [24]. The major roles of mTOR signaling in aging biology, including its involvement in cellular senescence, autophagy, metabolic regulation, and stem cell aging, are summarized in Table 1.
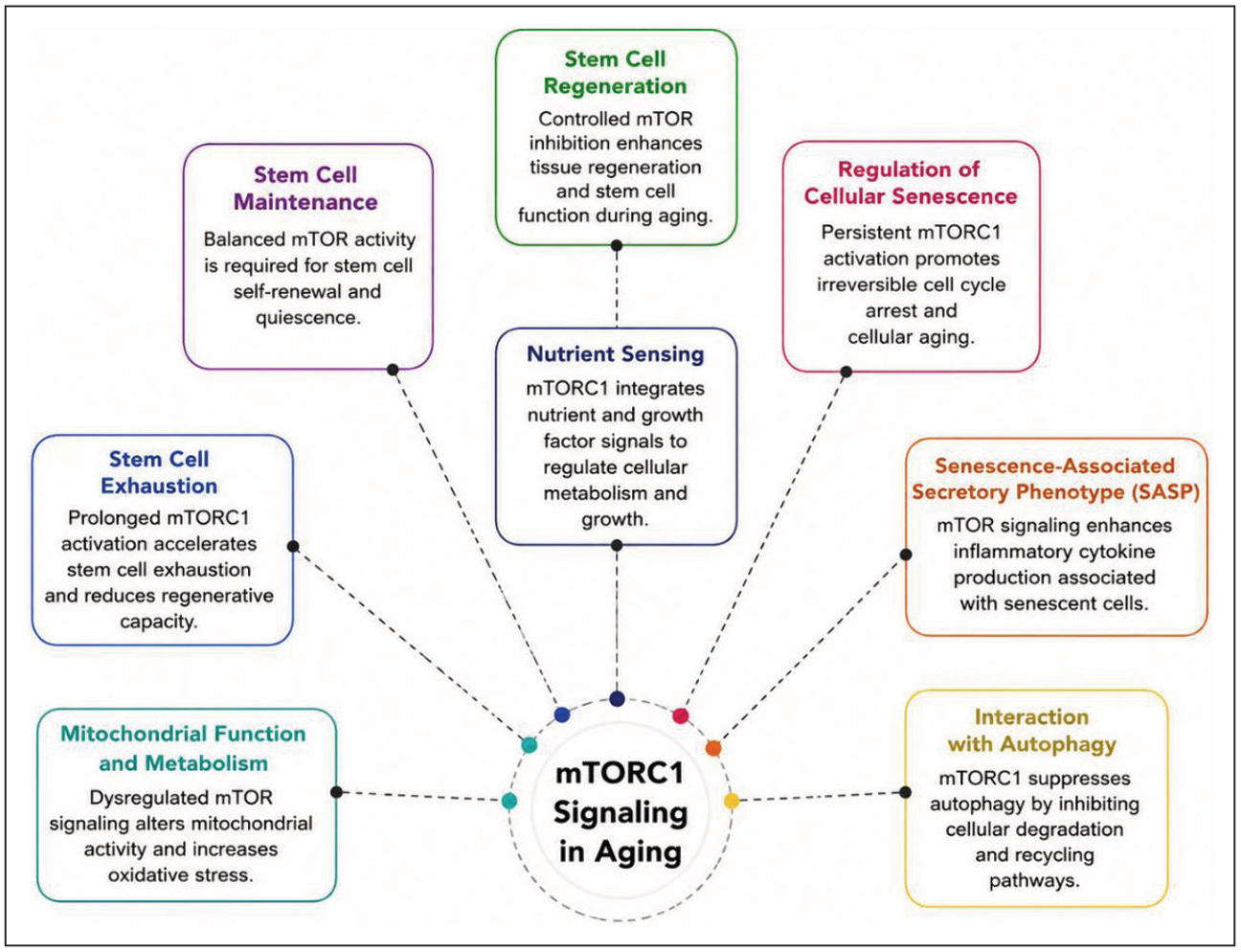
Figure 2. Role of mTORC1 signaling in aging. Dysregulated mTORC1 signaling contributes to multiple hallmarks of aging, including cellular senescence, senescence-associated secretory phenotype, impaired autophagy, mitochondrial dysfunction, altered nutrient sensing, and stem cell exhaustion, thereby promoting aging and age-related functional decline.
Table 1.
Role of mTOR signaling in aging biology.
| Section | Detailed mechanism | Impact on aging | References |
|---|---|---|---|
| mTOR and cellular senescence | |||
| Regulation of cellular senescence | Persistent activation of mTORC1 promotes cellular senescence by enhancing protein synthesis and metabolic activity while inhibiting autophagy. Excessive mTOR activity increases oxidative stress, DNA damage responses, and activation of cell cycle inhibitors such as p16 and p21, leading to irreversible cell cycle arrest. | Accelerates tissue aging and contributes to the accumulation of senescent cells in aging tissues. | [21–23] |
| SASP | mTOR signaling regulates the translation of inflammatory cytokines, chemokines, and growth factors that form the SASP. Activation of mTORC1 enhances production of pro-inflammatory molecules such as IL-6, IL-8, and matrix metalloproteinases. | SASP promotes chronic inflammation, tissue dysfunction, and age-related diseases such as cancer and neurodegeneration. | [22–24] |
| mTOR and autophagy | |||
| Interaction with autophagy | mTORC1 acts as a major negative regulator of Autophagy by phosphorylating ULK1 and preventing the initiation of autophagosome formation. Under nutrient-rich conditions, active mTORC1 suppresses autophagy, whereas nutrient deprivation inhibits mTORC1 and activates autophagic pathways. | Reduced autophagy contributes to accumulation of damaged proteins and organelles during aging. | [25] |
| Role in cellular homeostasis and longevity | Inhibition of mTOR signaling by compounds such as rapamycin promotes autophagy and enhances cellular stress resistance. This process facilitates the removal of dysfunctional mitochondria and aggregated proteins, thereby maintaining cellular homeostasis and extending lifespan in model organisms. | Improved longevity and delayed onset of age-related diseases. | [26] |
| mTOR and metabolic regulation | |||
| Nutrient sensing | mTOR functions as a central nutrient sensor that integrates signals from amino acids, glucose, oxygen, and growth factors. Activation of mTOR stimulates anabolic processes including lipid synthesis, nucleotide synthesis, and protein translation, enabling cells to adapt to nutrient availability. | Dysregulated nutrient sensing contributes to metabolic diseases and aging-related metabolic decline. | [27] |
| Mitochondrial function and metabolism | mTOR signaling influences mitochondrial biogenesis, oxidative phosphorylation, and metabolic reprogramming. Hyperactivation of mTOR can increase mitochondrial activity but also elevate reactive oxygen species production, leading to oxidative stress and cellular damage. | Alters energy metabolism and contributes to age-associated mitochondrial dysfunction. | [28] |
| mTOR and stem cell aging | |||
| Stem cell maintenance | Balanced mTOR signaling is essential for maintaining stem cell quiescence and self-renewal. Excessive activation of mTOR leads to stem cell exhaustion by promoting premature differentiation and metabolic stress. | Reduced regenerative capacity of tissues during aging. | [29] |
| Stem cell regeneration | Controlled inhibition of mTOR signaling has been shown to enhance stem cell function and tissue regeneration. For example, reduced mTOR activity improves hematopoietic and muscle stem cell maintenance, thereby supporting regenerative potential in aging tissues. | Potential therapeutic target for regenerative medicine and healthy aging. | [30] |
Notes: mTOR, mechanistic target of rapamycin; mTORC1, mechanistic target of rapamycin complex 1; SASP, senescence-associated secretory phenotype; IL-6, interleukin-6; IL-8, interleukin-8; ULK1, Unc-51-like kinase 1.
mTOR and cellular senescence
Cellular senescence is characterized by irreversible cell cycle arrest accompanied by metabolic and functional alterations. Persistent activation of mTORC1 promotes cellular senescence by enhancing protein synthesis and suppressing autophagy, leading to the accumulation of damaged proteins and organelles [31]. Hyperactivation of mTOR signaling also increases oxidative stress and activates cell cycle regulators such as p16 and p21, which contribute to senescence-associated growth arrest. Accumulation of senescent cells in tissues accelerates tissue dysfunction and aging-related degeneration [32].
SASP
Senescent cells actively secrete pro-inflammatory cytokines, chemokines, growth factors, and proteases collectively known as the SASP. mTORC1 regulates the translation and secretion of several SASP components, including interleukin-6 (IL-6), interleukin-8 (IL-8), and matrix metalloproteinases [33]. Chronic activation of SASP contributes to low-grade inflammation, tissue remodeling, and progression of age-related diseases such as cancer, neurodegeneration, and cardiovascular disorders [34].
mTOR and autophagy
Autophagy is an essential cellular degradation process responsible for the removal of damaged proteins and dysfunctional organelles. mTORC1 acts as a major negative regulator of autophagy by phosphorylating Unc-51-like kinase 1 (ULK1) and suppressing autophagosome formation under nutrient-rich conditions [35]. During nutrient deprivation or cellular stress, inhibition of the mTORC1 activates autophagy and promotes cellular recycling mechanisms. Age-associated decline in autophagy contributes to protein aggregation, mitochondrial dysfunction, and cellular damage, whereas pharmacological inhibition of mTOR enhances autophagy and supports cellular homeostasis and longevity [36].
mTOR and metabolic dysregulation
mTOR signaling functions as a central regulator of cellular metabolism by integrating nutrient and growth factor signals. Activation of mTOR promotes anabolic processes such as protein synthesis, lipid biosynthesis, and nucleotide production [37]. However, chronic mTOR activation contributes to metabolic imbalance, insulin resistance, and increased oxidative stress, all of which are associated with aging and age-related metabolic diseases [38]. Dysregulated mTOR signaling also affects mitochondrial activity and reactive oxygen species production, further accelerating cellular aging.
mTOR and stem cell exhaustion
Balanced mTOR activity is essential for maintaining stem cell quiescence, self-renewal, and regenerative capacity. Excessive activation of mTOR signaling drives premature stem cell differentiation and metabolic stress, leading to stem cell exhaustion during aging [39]. Reduced regenerative potential of stem cells contributes to impaired tissue repair and functional decline in aging organisms. Experimental studies have demonstrated that controlled inhibition of mTOR signaling can improve stem cell maintenance and enhance tissue regeneration, suggesting potential therapeutic applications in regenerative medicine and healthy aging [40].
mTOR signaling in age-related diseases
Hallmarks of aging and the role of mTORC1
Aging is a complex and progressive biological process characterized by gradual functional decline, accumulation of cellular and molecular damage, impaired regenerative capacity, metabolic dysregulation, and increased susceptibility to age-related diseases. To better understand the mechanisms underlying aging, several conserved hallmarks of aging have been identified, including genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, mitochondrial dysfunction, deregulated nutrient sensing, cellular senescence, stem cell exhaustion, impaired autophagy, and altered intercellular communication. These hallmarks interact with one another and collectively contribute to tissue degeneration and physiological decline during aging [11]. Among the major nutrient-sensing pathways, mTORC1 has emerged as a central regulator of aging and longevity. Dysregulated
or chronic activation of mTORC1 contributes to multiple hallmarks of aging through its effects on cellular growth, metabolism, protein synthesis, and stress responses. Persistent mTORC1 activation promotes excessive protein synthesis while suppressing autophagy, leading to loss of proteostasis and accumulation of damaged proteins and organelles. Inhibition of autophagy also impairs the clearance of dysfunctional mitochondria, thereby increasing oxidative stress and mitochondrial dysfunction [12].
Aberrant mTORC1 signaling further contributes to cellular senescence by enhancing metabolic activity, oxidative damage, and activation of senescence-associated pathways. Senescent cells subsequently release pro-inflammatory cytokines and mediators associated with the SASP, promoting chronic inflammation and tissue dysfunction. In addition, prolonged mTORC1 activation disrupts nutrient sensing and metabolic homeostasis, contributing to insulin resistance and age-related metabolic disorders. Excessive mTORC1 signaling also accelerates stem cell exhaustion by promoting premature differentiation and reducing self-renewal capacity, ultimately impairing tissue regeneration and repair [13]. Because mTORC1 integrates signals from nutrients, growth factors, and cellular energy status, it plays a critical role in coordinating anabolic and catabolic processes essential for cellular homeostasis. Therefore, pharmacological inhibition of mTORC1 has gained significant attention as a potential therapeutic strategy to delay aging, improve autophagy and metabolic balance, reduce chronic inflammation, and prevent the progression of age-associated diseases [41]. The major age-related diseases associated with dysregulated mTOR signaling are summarized in Figure 3.
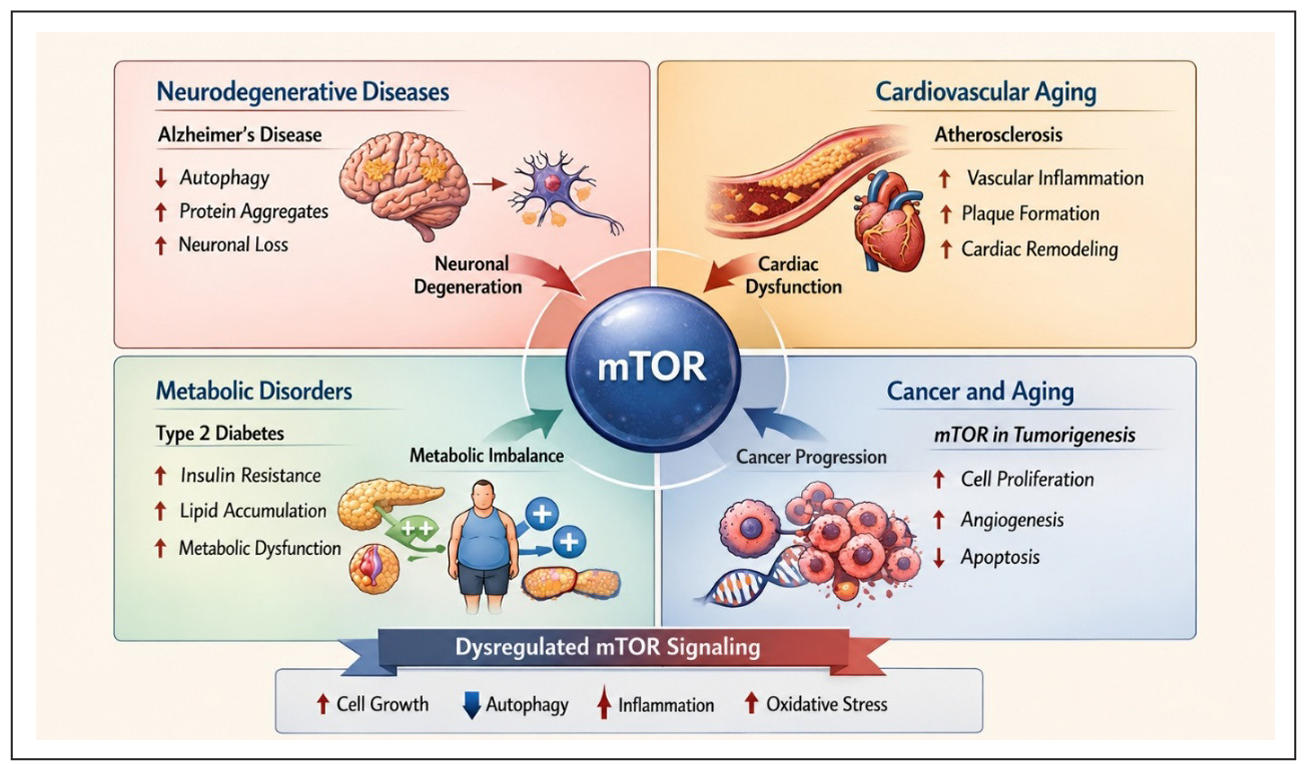
Figure 3. Dysregulated mTOR signaling in age-related diseases. Aberrant mTOR activity contributes to neurodegeneration, cardiovascular aging, metabolic disorders, and cancer progression by promoting inflammation, oxidative stress, cell proliferation, and reduced autophagy. Upward arrows (↑) indicate increased activity or pathological progression, whereas downward arrows (↓) indicate reduced activity or suppression of cellular processes.
Neurodegenerative diseases
Alzheimer’s disease
Amyloid-β plaque buildup, neurofibrillary tangles, and cognitive impairment are the hallmarks of Alzheimer’s disease, a progressive neurodegenerative illness. Because it affects autophagy, protein synthesis, and neuronal survival, dysregulation of the mTOR signaling pathway has been linked to the pathophysiology of Alzheimer’s disease. Amyloid-β and tau proteins can build up in neural tissues as a result of hyperactivation of mTOR signaling, which inhibits autophagic degradation mechanisms that remove harmful protein aggregates. Furthermore, increased mTOR activity accelerates neurodegeneration by causing neuroinflammation and synaptic dysfunction. Targeting mTOR may be a viable therapeutic approach for treating Alzheimer’s disease, as experimental studies have shown that inhibition of mTOR signaling can increase autophagy and decrease amyloid-β accumulation [41].
Parkinson’s disease
Another prevalent age-related neurodegenerative condition is Parkinson’s disease, which is defined by the formation of α-synuclein aggregates and the progressive death of dopaminergic neurons in the substantia nigra. In Parkinson’s disease, the mTOR Signaling pathway is crucial for controlling mitochondrial homeostasis, autophagy, and neuronal survival. Neuronal degeneration may result from impaired autophagic clearance of damaged mitochondria and aggregated proteins caused by dysregulated mTOR activation. Furthermore, oxidative stress and mitochondrial dysfunction—two major clinical aspects of Parkinson’s disease—have been connected to changes in mTOR signaling. Therefore, pharmacological manipulation of mTOR activity has been studied as a possible treatment strategy to halt the onset of neurodegenerative illness in Parkinson’s disease and restore cellular homeostasis [42].
Cardiovascular aging
Atherosclerosis
The buildup of lipid-rich plaques inside artery walls is known as atherosclerosis, and it is a significant factor in cardiovascular aging. Endothelial cell function, vascular smooth muscle cell proliferation, and inflammatory responses are among the biological processes implicated in atherosclerosis that are influenced by the mTOR signaling system. Plaque formation and progression are facilitated by overactivation of mTOR, which also causes vascular inflammation and aberrant smooth muscle cell proliferation. Moreover, oxidative stress and fat buildup in the vascular system may be exacerbated by mTOR-mediated metabolic changes. Therefore, it has been suggested that targeting mTOR signaling is a viable method for lowering vascular inflammation and stopping the formation of atherosclerotic lesions [43].
Cardiac hypertrophy
The heart’s structural and functional adaptation known as cardiac hypertrophy frequently happens as people age and in reaction to long-term stressors like high blood pressure. The regulation of cardiomyocyte growth and protein synthesis, two processes that lead to the development of cardiac hypertrophy, is largely dependent on mTOR signaling. Heart tissue undergoes pathological remodeling as a result of persistent stimulation of mTORC1, which promotes excessive protein synthesis and cellular expansion in cardiac muscle cells. In the end, this remodeling may worsen cardiac function and raise the risk of heart failure in older people. Pharmacological suppression of mTOR signaling has been shown in experiments to reduce ventricular hypertrophy and enhance cardiac function, underscoring its therapeutic potential in aging-related cardiovascular disorders [44].
Metabolic disorders
Type 2 diabetes
Insulin resistance and compromised glucose homeostasis are hallmarks of type 2 diabetes, a chronic metabolic disease. mTOR signaling is crucial for controlling metabolic balance and insulin signaling pathways. Insulin resistance may result from long-term mTORC1 activity, which phosphorylates insulin receptor substrate proteins and inhibits insulin receptor signaling. Furthermore, dysregulated mTOR activity impacts glucose metabolism and pancreatic β-cell function, aggravating hyperglycemia. As a result, mTOR signaling modification has been investigated as a possible treatment strategy for enhancing insulin sensitivity and controlling metabolic issues related to type 2 diabetes [45].
Obesity and insulin resistance
Insulin resistance and obesity are closely related metabolic disorders that have a strong correlation with lifestyle variables and aging. The mTOR pathway influences the development of obesity by controlling energy balance, lipid metabolism, and adipocyte differentiation. Adipogenesis and lipid buildup in adipose tissues can be encouraged by excessive food intake and long-term mTOR signaling activation. Persistent mTOR activation also adds to systemic insulin resistance by interfering with insulin signaling pathways. The significance of balanced mTOR activity in preserving metabolic health is highlighted by the fact that these metabolic changes raise the risk of obesity-related comorbidities, such as cardiovascular disease and type 2 diabetes [46].
Cancer and aging
Age-related increases in cancer incidence are substantial, and dysregulated mTOR signaling has been found to be a key factor in the growth and spread of tumors. Essential biological functions like protein synthesis, cell proliferation, metabolism, and survival are all regulated by the mTOR pathway and are often disrupted in cancer cells. Tumorigenesis and the advancement of cancer are facilitated by hyperactivation of mTOR signaling, which encourages unchecked cell proliferation and metabolic reprogramming. Furthermore, mTOR signaling in cancer cells can be enhanced by mutations or abnormal activation of upstream regulators like PI3K and AKT. Due to its crucial involvement in tumor biology, mTOR has emerged as a significant therapeutic target in oncology, and a number of mTOR inhibitors are either being utilized or being researched for the treatment of various cancer types [47].
Therapeutic targeting of mTOR for longevity
One of the most promising therapeutic targets for interventions meant to increase longevity and improve healthspan is the mTOR pathway. Modulation of this system can have a substantial impact on metabolic regulation, autophagy, and cellular stress responses because mTOR integrates information relating to nutrition availability, growth factors, and cellular energy status. Inhibition of mTOR signaling has been shown in numerous experimental studies in model organisms, such as yeast, worms, flies, and mammals, to postpone aging processes and lower the prevalence of age-related illnesses. By inhibiting excessive mTOR activity and encouraging cellular maintenance mechanisms like autophagy, pharmacological treatments like rapamycin and its derivatives, also referred to as rapalogs, have been demonstrated to increase longevity and improve physiological performance [48]. Figure 4 illustrates the mechanism of action of major mTOR inhibitors and their downstream effects on autophagy, protein synthesis, cellular senescence, and longevity-associated pathways.
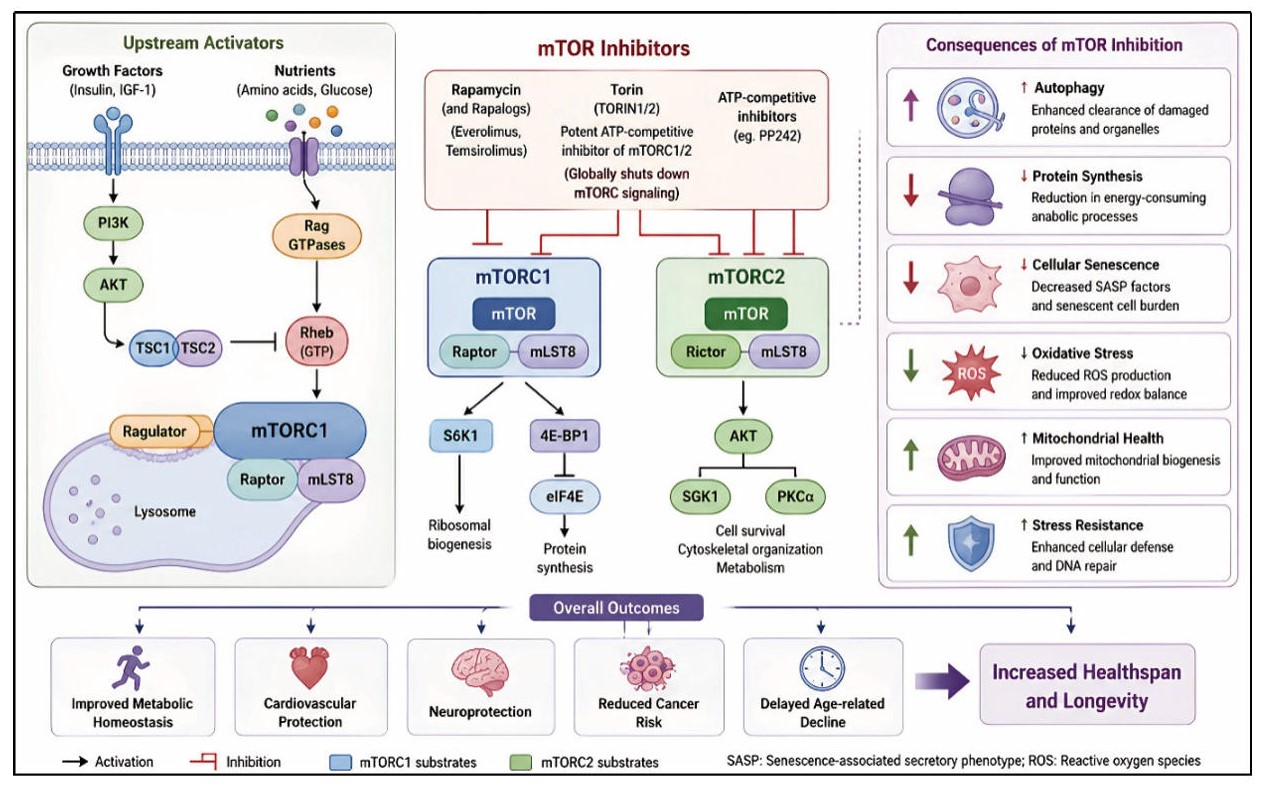
Figure 4. Mechanism and biological effects of mTOR inhibitors. Rapamycin, rapalogs, and Torin inhibit mTOR signaling, promoting autophagy, reducing protein synthesis and cellular senescence, and improving metabolic homeostasis and longevity.
Apart from pharmaceutical inhibitors, dietary and lifestyle modifications like calorie restriction and intermittent fasting can also affect mTOR activity and promote longevity. These treatments improve metabolic efficiency and cellular repair processes by decreasing nutrition signaling and activating cellular stress resistance pathways [12]. The major therapeutic strategies targeting mTOR signaling for longevity and healthy aging are summarized in Table 2. As a result, therapeutic targeting of the mTOR pathway is becoming more widely acknowledged as a viable technique for avoiding or postponing a number of age-related ailments, including cancer, metabolic dysfunction, neurodegeneration, and cardiovascular diseases. Therefore, the goal of ongoing research is to create safer and more effective mTOR-targeted treatments that can encourage healthy aging without causing serious side effects [49].
Table 2.
Therapeutic strategies targeting mTOR signaling for longevity and healthy aging, including pharmacological inhibitors, natural compounds,
and lifestyle interventions that regulate autophagy, metabolism, and cellular aging.
| Section | Intervention | Mechanism on mTOR | Potential benefits | References |
|---|---|---|---|---|
| Pharmacological inhibitors | Rapamycin | Directly inhibits mTORC1 and promotes autophagy | Extends lifespan and delays age-related diseases in experimental models | [50] |
| Everolimus | Rapamycin analog that suppresses mTORC1 signaling and cell growth | Improves immune and metabolic regulation | [51] | |
| Temsirolimus | Inhibits mTOR-mediated cell proliferation and angiogenesis | Reduces pathological cell growth and tumor development | [52] | |
| Torin | ATP-competitive inhibitor that globally suppresses mTORC1/2 signaling | Promotes autophagy, suppresses cellular senescence, and shows potential anti-aging effects | [53] | |
| Natural compounds modulating mTOR | Resveratrol | Activates AMPK and indirectly suppresses mTOR signaling | Enhances mitochondrial function and longevity pathways | [54] |
| Curcumin | Inhibits PI3K/AKT/mTOR signaling and reduces inflammation | Provides antioxidant and anti-aging effects | [55] | |
| Polyphenols and dietary phytochemicals | Plant-derived compounds regulate nutrient-sensing pathways including mTOR | Support metabolic balance and cellular protection | [56] | |
| Lifestyle interventions | Caloric restriction | Reduces nutrient signaling and suppresses mTOR activity | Improves metabolic health and lifespan | [57] |
| Intermittent fasting | Periodic fasting inhibits mTOR and activates autophagy | Enhances stress resistance and longevity | [58] | |
| Physical exercise | Balances AMPK–mTOR signaling and improves metabolism | Promotes cardiovascular and metabolic health | [59] |
Emerging anti-aging strategies
Combination therapies targeting mTOR and other pathways
According to recent studies, anti-aging treatments that target the mTOR pathway in conjunction with other longevity-related signaling networks may be more successful. A number of interrelated mechanisms, including AMPK, insulin/IGF-1 signaling, and sirtuin-mediated metabolic control, are involved in the intricate and multifaceted process of aging. Therefore, treatment approaches that combine mTOR inhibitors with these pathway modulators may improve metabolic balance, increase longevity, and strengthen cellular stress resistance. In addition to attaining wider protective effects against age-related functional decline and disease development, combination treatments may lessen the possible adverse consequences linked to long-term blockage of a specific mechanism [60].
Integration with senolytics
Combining mTOR-targeted treatments with senolytic agents—which specifically destroy senescent cells—is another potential approach in the study of aging. As people age, senescent cells proliferate and emit pro-inflammatory substances known as the SASP, which lead to tissue dysfunction. While senolytic substances eliminate already-accumulated senescent cells from tissues, inhibition of mTOR signaling can prevent the onset of cellular senescence and lower inflammatory signaling. Therefore, by avoiding senescence and eliminating senescent cell populations, the combination use of mTOR modulators and senolytics may offer synergistic effects that improve tissue function and encourage healthy aging [61].
Precision medicine approaches for aging interventions
Advancements in molecular profiling and precision medicine have created new opportunities for the development of personalized anti-aging therapies. The manner in which the mTOR pathway contributes to aging and disease risk can be greatly influenced by individual differences in genetic background, metabolic status, and environmental exposure. By identifying particular molecular signatures and biomarkers linked to dysregulated mTOR signaling, precision medicine techniques enable patient-specific therapies. Researchers can create tailored treatment plans that maximize mTOR activity modulation while reducing side effects by combining genetic, proteomic, and metabolic data [62]. This will ultimately increase the efficacy of aging-related therapies and prolong healthy lifespan. Figure 5 illustrates the major emerging anti-aging strategies targeting mTOR signaling, including combination therapies, senolytic integration, and precision medicine approaches.
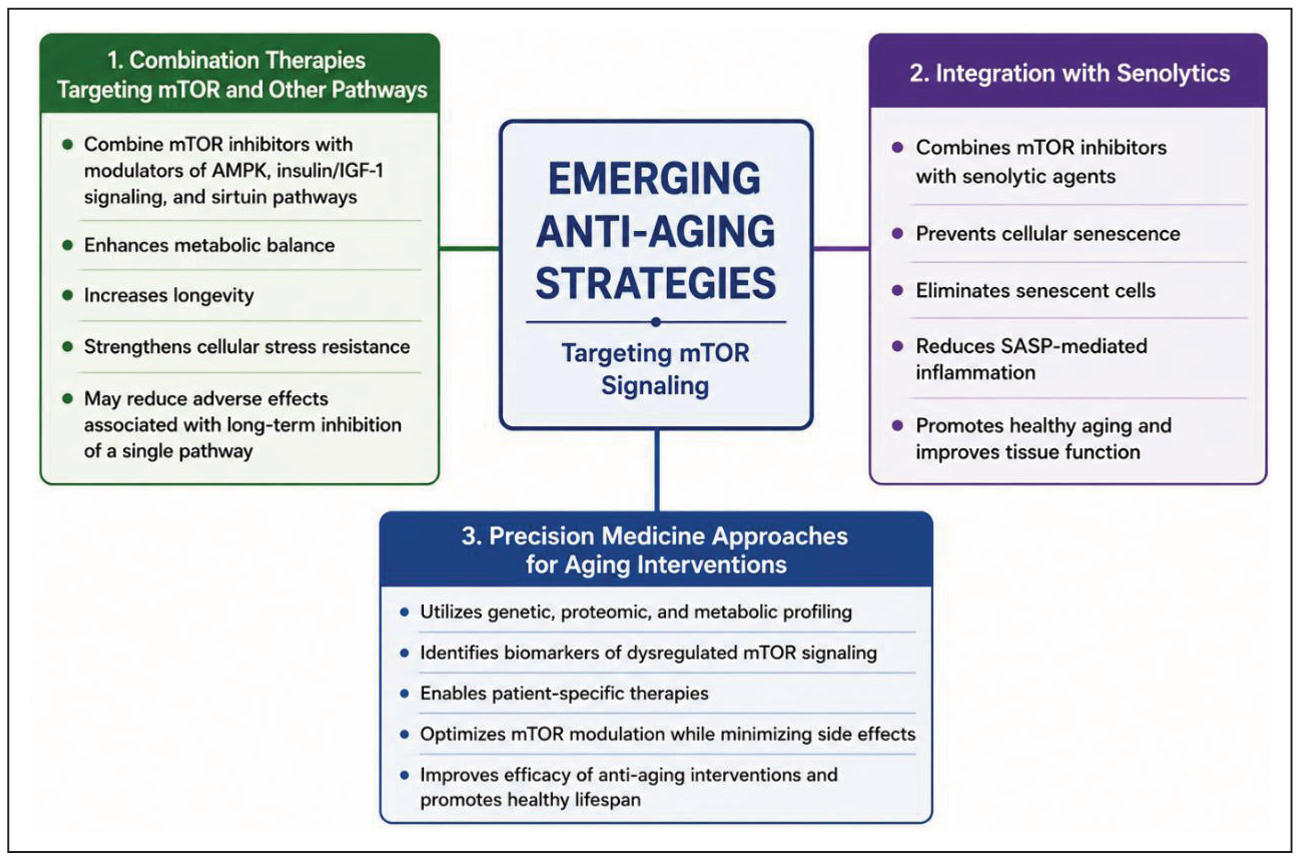
Figure 5. Emerging anti-aging strategies targeting mTOR signaling. The figure summarizes major therapeutic approaches, including combination therapies targeting mTOR and related pathways, integration of mTOR inhibitors with senolytic agents, and precision medicine approaches for personalized aging interventions.
Regulation of mTORC1 by long non-coding RNAs (lncRNAs)
LncRNAs have emerged as important regulators of mTORC1 signaling pathway and are increasingly recognized for their role in aging and age-related diseases. lncRNAs regulate mTORC1 activity through diverse mechanisms, including modulation of upstream signaling molecules such as PI3K/AKT, AMPK, and growth factor receptors, as well as through epigenetic and post-transcriptional regulation of mTOR-associated genes. Certain lncRNAs promote mTORC1 activation, leading to enhanced protein synthesis, reduced autophagy, cellular senescence, and metabolic dysregulation, whereas others suppress mTOR signaling and exert protective effects against aging-related cellular damage [63].
Recent studies suggest that dysregulation of the lncRNA–mTORC1 axis contributes to several pathological conditions associated with aging, including cancer, neurodegenerative disorders, metabolic dysfunction, and cardiovascular diseases. Furthermore, targeting lncRNA-mediated regulation of mTORC1 has emerged as a promising therapeutic strategy for modulating cellular senescence, improving autophagy, and promoting healthy aging. Therefore, understanding the complex interaction between lncRNAs and mTORC1 signaling may provide new insights into the molecular mechanisms of aging and facilitate the development of novel anti-aging interventions [64].
Challenges
Limitations of long-term mTOR inhibition
Long-term suppression of mTOR signaling may have unanticipated physiological repercussions, even though inhibition of the mTOR pathway has shown encouraging outcomes in prolonging longevity and increasing healthspan in various experimental models. Since mTOR controls vital physiological functions like protein synthesis, cell proliferation, and metabolic homeostasis, long-term suppression may disrupt normal cellular processes and tissue healing systems. In order to minimize negative effects, it is crucial to optimize therapeutic options such as intermittent dosing or selective targeting of particular mTOR complexes. Continuous reduction of mTOR activity may potentially interfere with adaptive stress responses and metabolic regulation [65].
Side effects and immunosuppression
Because of its immunosuppressive and antiproliferative qualities, pharmacological inhibitors of mTOR, such as rapamycin and its variants, are frequently employed in cancer treatment and organ transplantation. Long-term use of these drugs, however, is linked to a number of negative consequences, including insulin resistance, immunosuppression, metabolic dysregulation, delayed wound healing, and heightened vulnerability to infections. The need for safer mTOR-targeted medicines with better selectivity and tolerance is highlighted by these adverse effects, which are especially worrisome in older people who may already be experiencing age-related immunological loss [66].
Need for clinical trials targeting aging rather than disease
Instead of addressing aging itself, the majority of mTOR inhibitor clinical trials have concentrated on treating particular diseases like cancer, cardiovascular conditions, or metabolic diseases. Interventions targeted at decreasing the biological aging process may be able to prevent or postpone many diseases at once because aging is the primary risk factor underlying a number of chronic ailments. However, extended research periods, a lack of standardized biomarkers of aging, and regulatory constraints continue to make it difficult to design clinical trials that directly assess aging. The advancement of clinical research on longevity therapies will depend on the establishment of validated endpoints and aging biomarkers [67].
Future perspectives
It is anticipated that future studies on mTOR-based anti-aging treatments will concentrate on creating safer and more accurate treatment approaches that alter the rapamycin pathway’s molecular target without producing systemic toxicity. The discovery of trustworthy biomarkers that represent mTOR activity and biological aging may be made possible by developments in systems biology, genomics, proteomics, and metabolomics. These biomarkers have the potential to monitor therapy responses, optimize dosage schedules, and tailor therapies based on individual biological profiles. Additionally, new strategies—including combination therapies that target several longevity pathways, intermittent drug delivery, and specific inhibition of mTORC1—may improve the efficacy of aging therapy. To confirm the safety and long-term advantages of mTOR-targeted treatments for encouraging healthy aging in humans, more translational research and well-planned clinical trials will be necessary [68].
Conclusions
By combining information about nutrition availability,
growth factors, and cellular energy state, the mTOR signaling pathway has become a
key regulator of aging and lifespan. Cellular senescence,
defective autophagy, metabolic imbalance, and stem cell failure are among the molecular processes linked to aging that are influenced by
dysregulation of mTOR signaling. Age-related illnesses like cancer, metabolic syndromes, cardiovascular
disease, and neurological disorders are mostly caused by these changes. As a result, mTOR pathway
modification has drawn a lot of interest as a potentially effective treatment approach for prolonging
life expectancy and postponing the
development of age-related diseases [69].
In experimental models, pharmacological inhibitors like rapamycin and its analogues, as well as natural
substances and lifestyle modifications like exercise and calorie restriction, have shown promise in
controlling mTOR activity and extending life. Furthermore, new approaches to the development of more
potent anti-aging medications include combination therapy, senolytic methods, and precision
medicine-based interventions. Despite these developments, there are still a number of issues,
such as immunosuppressive effects, long-term safety concerns, and the requirement for carefully
planned clinical trials that focus on aging rather than specific diseases. To translate these
findings into safe and efficient treatments that support healthy aging in humans, future studies
concentrating on specific mTOR regulation and the discovery of
trustworthy biomarkers will be essential [70].
Declarations
Author contributions
Made substantial contributions to conception and design of the study, as well as investigation and writing of the original draft: Rasal PB, Kasar GN. Performed data curation, visualization, and formal analysis, as well as provided resources: Rasal PB, Kasar GN, Upaganlawar AB, Mahajan MS.
Conflict of interest
The authors confirm that there are no known conflicts of interest.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
AI and AI-assisted tools statement
No AI tools were used in the preparation of this article.
References
1. Beard J, Officer A, de Carvalho I, Sadana R, Pot A, Michel J, et al. The world report on ageing and health: a policy framework for healthy ageing. Lancet, 2016, 387(10033): 2145-2154. [Crossref]
2. López-Otín C, Blasco M, Partridge L, Serrano M, & Kroemer G. The hallmarks of aging. Cell, 2013, 153(6): 1194-1217. [Crossref]
3. Fekete M, Major D, Feher A, Fazekas-Pongor V, & Lehoczki A. Geroscience and pathology: a new frontier in understanding age-related diseases. Pathol Oncol Res, 2024, 30: 1611623. [Crossref]
4. López-Otín C, Blasco M, Partridge L, Serrano M, & Kroemer G. Hallmarks of aging: an expanding universe. Cell, 2023, 186(2): 243-278. [Crossref]
5. Campisi J, Kapahi P, Lithgow G, Melov S, Newman J, & Verdin E. From discoveries in ageing research to therapeutics for healthy ageing. Nature, 2019, 571(7764): 183-192. [Crossref]
6. Saxton R, & Sabatini D. mTOR signaling in growth, metabolism, and disease. Cell, 2017, 168(6): 960-976. [Crossref]
7. Marafie S, Al-Mulla F, & Abubaker J. mTOR: its critical role in metabolic diseases, cancer, and the aging process. Int J Mol Sci, 2024, 25(11): 6141-6152. [Crossref]
8. Lamming D. Inhibition of the mechanistic target of rapamycin (mTOR)-rapamycin and beyond. Cold Spring Harb Perspect Med, 2016, 6(5): a025924. [Crossref]
9. Gargalionis A, Papavassiliou K, & Papavassiliou A. mTOR signaling: recent progress. Int J Mol Sci, 2024, 25(5): 2587-2598. [Crossref]
10. Liu G, & Sabatini D. mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol, 2020, 21(4): 183-203. [Crossref]
11. Kim J, & Guan K. mTOR as a central hub of nutrient signalling and cell growth. Nat Cell Biol, 2019, 21(1): 63-71. [Crossref]
12. Jhanwar-Uniyal M, Zeller S, Spirollari E, Das M, Hanft S, & Gandhi C. Discrete mechanistic target of rapamycin signaling pathways, stem cells, and therapeutic targets. Cells, 2024, 13(5): 409-421. [Crossref]
13. Sorrenti V, Benedetti F, Buriani A, Fortinguerra S, Caudullo G, Davinelli S, et al. Immunomodulatory and antiaging mechanisms of resveratrol, rapamycin, and metformin: focus on mTOR and AMPK signaling networks. Pharmaceuticals, 2022, 15(8): 912-924. [Crossref]
14. Fu W, & Hall M. Regulation of mTORC2 signaling. Genes, 2020, 11(9): 1045-1056. [Crossref]
15. Surugiu R, Iancu M, Vintilescu ŞB, Stepam M, Burduşel D, Genunche-Dumitrescu A, et al. Molecular mechanisms of healthy aging: the role of caloric restriction, intermittent fasting, Mediterranean diet, and ketogenic diet—a scoping review. Nutrients, 2024, 16(17): 2878-2890. [Crossref]
16. Sparta B, Kosaisawe N, Pargett M, Patankar M, DeCuzzi N, & Albeck J. Continuous sensing of nutrients and growth factors by the mTORC1-TFEB axis. Elife, 2023, 12: e74903. [Crossref]
17. Hardie D, & Ashford M. AMPK: regulating energy balance at the cellular and whole body levels. Physiology (Bethesda), 2014, 29(2): 99-107. [Crossref]
18. Laplante M, & Sabatini D. mTOR signaling in growth control and disease. Cell, 2012, 149(2): 274-293. [Crossref]
19. Magnuson B, Ekim B, & Fingar D. Regulation and function of ribosomal protein S6 kinase (S6K) within mTOR signalling networks. Biochem J, 2012, 441(1): 1-21. [Crossref]
20. Hay N, & Sonenberg N. Upstream and downstream of mTOR. Genes Dev, 2004, 18(16): 1926-1945. [Crossref]
21. Wang J, Eming S, & Ding X. Role of mTOR signaling cascade in epidermal morphogenesis and skin barrier formation. Biology, 2022, 11(6): 931-942. [Crossref]
22. Blagoslovnny M. Cellular senescence: when growth stimulation meets cell cycle arrest. Aging, 2023, 15(4): 905-913. [Crossref]
23. Chouhan S, Kumar A, Piprode V, Dasgupta A, Singh S, & Khalique A. Regulatory-associated protein of mTOR-mediated signaling: a nexus between tumorigenesis and disease. Targets, 2024, 2(4): 341-371. [Crossref]
24. Cayo A, Segovia R, Venturini W, Moore-Carrasco R, Valenzuela C, & Brown N. mTOR activity and autophagy in senescent cells, a complex partnership. Int J Mol Sci, 2021, 22(15): 8149-8163. [Crossref]
25. Ebata H, & Hansen M. Links between autophagy and healthy aging. J Mol Biol, 2026, 438(6): 169656. [Crossref]
26. Jiao Y, Li G, Li Q, Ali R, Qin L, Li W, et al. mTOR (mechanistic target of rapamycin) inhibition decreases mechanosignaling, collagen accumulation, and stiffening of the thoracic aorta in elastin-deficient mice. Arterioscler Thromb Vasc Biol, 2017, 37(9): 1657-1666. [Crossref]
27. Kennedy B, Berger S, Brunet A, Campisi J, Cuervo A, Epel E, et al. Geroscience: linking aging to chronic disease. Cell, 2014, 159(4): 709-713. [Crossref]
28. Ruggiero C, Tafaro L, Cianferotti L, Tramontana F, Macchione IG, Caffarelli C, et al. Targeting the hallmarks of aging with vitamin D: starting to decode the myth. Nutrients, 2024, 16(6): 906-918. [Crossref]
29. Wei X, Luo L, & Chen J. Roles of mTOR signaling in tissue regeneration. Cells, 2019, 8(9): 1075-1086. [Crossref]
30. Neves J, Demaria M, Campisi J, & Jasper H. Of flies, mice, and men: evolutionarily conserved tissue damage responses and aging. Dev Cell, 2015, 32(1): 9-18. [Crossref]
31. Demidenko Z, & Blagoslovnny M. Growth stimulation leads to cellular senescence when the cell cycle is blocked. Cell Cycle, 2008, 7(21): 3355-3361. [Crossref]
32. Herranz N, & Gil J. Mechanisms and functions of cellular senescence. J Clin Invest, 2018, 128(4): 1238-1246. [Crossref]
33. Laberge R, Sun Y, Orjala A, Patil C, Freund A, Zhou L, et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol, 2015, 17(8): 1049-1061. [Crossref]
34. Coppé J, Desprez P, Krtolica A, & Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol, 2010, 5: 99-118. [Crossref]
35. Kim J, Kundu M, Viollet B, & Guan K. AMPK and mTOR regulate autophagy through direct phosphorylation of ULK1. Nat Cell Biol, 2011, 13(2): 132-141. [Crossref]
36. Rubinsztein D, Mariño G, & Kroemer G. Autophagy and aging. Cell, 2011, 146(5): 682-695. [Crossref]
37. Shimobayashi M, & Hall M. Making new contacts: the mTOR network in metabolism and signalling crosstalk. Nat Rev Mol Cell Biol, 2014, 15(3): 155-162. [Crossref]
38. Lamming D, & Sabatini D. A central role for mTOR in lipid homeostasis. Cell Metab, 2013, 18(4): 465-469. [Crossref]
39. Yilmaz Ö, Katajisto P, Lamming D, Gültekin Y, Bauer-Rowe K, Sengupta S, et al. mTORC1 in the Paneth cell niche couples intestinal stem-cell function to calorie intake. Nature, 2012, 486(7404): 490-495. [Crossref]
40. Garcia-Prat L, Martinez-Vicente M, Perdiguero E, Ortet L, Rodríguez-Ubreva J, Rebollo E, et al. Autophagy maintains stemness by preventing senescence. Nature, 2016, 529(7584): 37-42. [Crossref]
41. Davoody S, Asgari Taei A, Khodabakhsh P, & Dargahi L. mTOR signaling and Alzheimer's disease: what we know and where we are? CNS Neurosci Ther, 2024, 30(4): e14463. [Crossref]
42. Zhu Z, Yang C, Iyaswamy A, Krishnamoorthi S, Sreenivasmurthy S, Liu J, et al. Balancing mTOR signaling and autophagy in the treatment of Parkinson's disease. Int J Mol Sci, 2019, 20(3): 728-739. [Crossref]
43. Liu Z, Fan Y, Zhang Z, Fang Y, Cheng X, Yang Q, et al. mTOR in the mechanisms of atherosclerosis and cardiovascular disease. Discov Med, 2021, 31(164): 129-140.
44. Wang J, Huang Y, Wang Z, Liu J, Liu Z, Yang J, et al. The mTOR signaling pathway: key regulator and therapeutic target for heart disease. Biomedicines, 2025, 13(2): 397-410. [Crossref]
45. Stanciu S, Jinga M, Miricescu D, Stefani C, Nica R, Stanescu S, et al. mTOR dysregulation, insulin resistance, and hypertension. Biomedicines, 2024, 12(8): 1802-1815. [Crossref]
46. Huang X, Liu G, Guo J, & Su Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int J Biol Sci, 2018, 14(11): 1483-1496. [Crossref]
47. Bazzichetto C, Conciatori F, Falcone I, & Ciuffreda L. Translational landscape of mTOR signaling in integrating cues between cancer and tumor microenvironment. Adv Exp Med Biol, 2020, 1223: 69-80. [Crossref]
48. Blagosklonny M. Rapamycin for longevity: opinion article. Aging, 2019, 11(19): 8048-8067. [Crossref]
49. Farrelly C. "Post-protean" public health and the geroscience hypothesis. Aging Dis, 2024, 15(2): 449-458. [Crossref]
50. Roark K, & Ifland P. Rapamycin for longevity: the pros, the cons, and future perspectives. Front Aging, 2025, 6: 1628187. [Crossref]
51. Mannick J, Morris M, Hockey H, Roma G, Beibel M, Kulmatycki K, et al. TORC1 inhibition enhances immune function and reduces infections in the elderly. Sci Transl Med, 2018, 10(449): e3003794. [Crossref]
52. Popova N, & Jücker M. The role of mTOR signaling as a therapeutic target in cancer. Int J Mol Sci, 2021, 22(4): 1743-1756. [Crossref]
53. Thoreen C, Kang S, Chang J, Liu Q, Zhang J, Gao Y, et al. An ATP-competitive mammalian target of rapamycin inhibitor reveals rapamycin-resistant functions of mTORC1. J Biol Chem, 2009, 284(12): 8023-8032. [Crossref]
54. Koç T, Doğan Ş, & Karadayın M. Potential using of resveratrol and its derivatives in medicine. Eurasian J Med, 2024, 56(2): 136-141. [Crossref]
55. Izadi M, Sadri N, Abdi A, Zadeh M, Jalaei D, Ghazimoradi M, et al. Longevity and anti-aging effects of curcumin supplementation. Geroscience, 2024, 46(3): 2933-2950. [Crossref]
56. Panzella L. Polyphenols and their impact on human health. Int J Mol Sci, 2023, 24(23): 16683. [Crossref]
57. Xia S, Wang M, Mo X, Wang J, Zheng S, & Shen X. Moderate dietary restriction across generations promotes sustained health and extends lifespan by enhancing antioxidant capacity in Bombyx mori. Sci Rep, 2025, 15(1): 17533. [Crossref]
58. Adafer R, Messaadi W, Meddahi M, Patey A, Haderbache A, Bayen S, et al. Food timing, circadian rhythm and chrononutrition: a systematic review of time-restricted eating's effects on human health. Nutrients, 2020, 12(12): 3770-3784. [Crossref]
59. Furrer R, Hawley J, & Handschin C. The molecular athlete: exercise physiology from mechanisms to medals. Physiol Rev, 2023, 103(3): 1693-1787. [Crossref]
60. Partridge L, Fuentealba M, & Kennedy B. The quest to slow ageing through drug discovery. Nat Rev Drug Discov, 2020, 19(8): 513-532. [Crossref]
61. Benhamú B, Martín-Fontecha M, Vázquez-Villa H, López-Rodríguez M, & Ortega-Gutiérrez S. New trends in aging drug discovery. Biomedicines, 2022, 10(8): 2006-2017. [Crossref]
62. Rosenfeld M, & Ladiges W. Pharmaceutical interventions to slow human aging. Are we ready for cocktails? Aging Pathobiol Ther, 2022, 4(2): 51-52. [Crossref]
63. Abouheiden K. Regulation of mTOR signaling by long non-coding RNA. Biochim Biophys Acta Gene Regul Mech, 2020, 1863(4): 194449. [Crossref]
64. Wang P, Xue Y, Han Y, Lin L, Wu C, Xu S, et al. The STAT3-binding long noncoding RNA lnc-DC controls human dendritic cell differentiation. Science, 2014, 344(6181): 310-313. [Crossref]
65. Blagosklonny M. From rapalogs to anti-aging formula. Oncotarget, 2017, 8(22): 35492-35507. [Crossref]
66. Zhang P, Catterson J, Grönke S, & Partridge L. Inhibition of S6K lowers age-related inflammation and increases lifespan through the endolysosomal system. Nat Aging, 2024, 4(4): 491-509. [Crossref]
67. Forman D, Kuchel G, Newman J, Kirkland J, Volpi E, Taffet G, et al. Impact of geroscience on therapeutic strategies for older adults with cardiovascular disease: JACC scientific statement. J Am Coll Cardiol, 2023, 82(7): 631-647. [Crossref]
68. Raghuvanshi K, Raghuvanshi D, Kumar D, Nepovimova E, Valko M, Kuca K, et al. Exploring the role of mTOR pathway in aging and age-related disorders. EXCLI J, 2025, 24: 992-1015. [Crossref]
69. Vitto V, Bianchin S, Zolondick A, Pellielo G, Rimessi A, Chianese D, et al. Molecular mechanisms of autophagy in cancer development, progression, and therapy. Biomedicines, 2022, 10(7): 1596-1609. [Crossref]
70. Chrienova Z, Nepovimova E, & Kuca K. The role of mTOR in age-related diseases. J Enzyme Inhib Med Chem, 2021, 36(1): 1679-1693. [Crossref]