Open Access | Editorial
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Brain iron in ageing: pixels to pathways
* Corresponding author: Thomas Welton
Mailing address: Level 6, National Neuroscience Institute, 11 Jln
Tan Tock Seng, Singapore.
Email: thomas_welton@nni.com.sg
This article belongs to the Special Issue: Ever evolving hallmarks of aging
Received: 15 May 2026 / Accepted: 19 May 2026 / Published: 30 June 2026
DOI: 10.31491/APT.2026.06.214
Abstract
Nigrostriatal iron deposition is a reproducible feature of brain ageing and Parkinsonian neurodegeneration, but its biological and therapeutic significance remains incompletely understood. Iron-sensitive MRI methods, including quantitative susceptibility mapping (QSM) and related approaches, can localize regional susceptibility changes in the substantia nigra and striatum in vivo, yet imaging alone cannot distinguish whether these changes reflect adaptive iron storage, impaired trafficking, mitochondrial stress, inflammatory activation, neuromelanin-associated iron, ferroptosis-like vulnerability, or downstream neurodegeneration. Despite recent emphasis on metabolic dysfunction, inflammaging, proteostasis failure, mitophagy, and other ageing pathways, iron biology intersects with these processes in ways that are still poorly resolved at the molecular level. This editorial argues that nigrostriatal iron-sensitive MRI should be reframed as a molecularly informative endophenotype. Integrating it with population-scale genomic, transcriptomic and multi-omic data offers a route to identify the genes, pathways and cell types regulating iron accumulation, distinguish physiological ageing from pathological dysregulation, prioritize therapeutic targets and support biomarker-guided intervention strategies in aging-related neurodegenerative disease.
Keywords
Nigrostriatal iron, ageing, Parkinson’s disease, post-GWAS, omics, therapeutic target discovery
Nigrostriatal iron accumulation is a consistent feature of brain ageing and Parkinsonian neurodegeneration, but its biological meaning remains uncertain. Recent studies have emphasized several recurring themes in ageing biology, including metabolic dysfunction, chronic inflammation, proteostasis failure, mitochondrial impairment, mitophagy and therapeutic modulation of age-related pathways [1-4]. Brain iron is essential for mitochondrial respiration, myelin maintenance and neurotransmitter synthesis, yet poorly regulated iron can drive oxidative stress, lipid peroxidation and cellular vulnerability. The substantia nigra and striatum are particularly relevant given their central roles in dopaminergic signaling and their well-documented age- and disease-related changes on iron-sensitive imaging. This perspective argues that integrating iron-sensitive MRI with multi-omic data offers a powerful route to move beyond correlative descriptions and identify the specific molecular programmes regulating iron handling in ageing and disease (Figure 1).
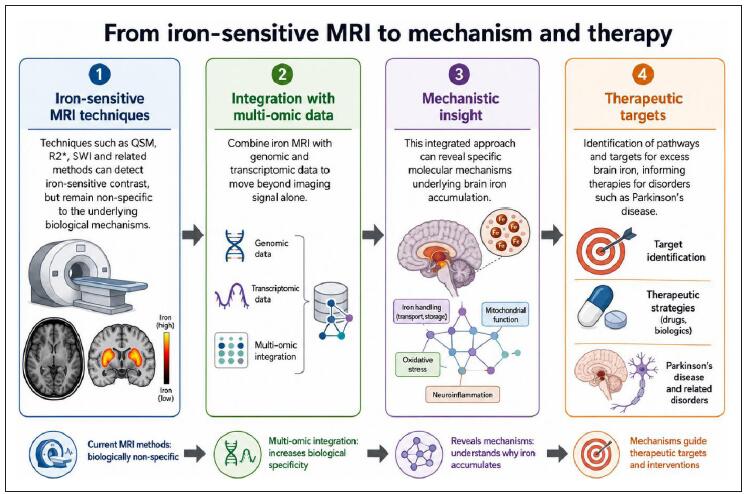
Figure 1. Integrating iron-sensitive MRI with multi-omic data to identify mechanisms and therapeutic targets.
Iron-sensitive MRI techniques, including quantitative susceptibility mapping, R2*, susceptibility-weighted imaging and related methods, can detect brain regions with altered iron-sensitive contrast, but these signals are not specific to the underlying biological processes that drive iron accumulation. Integrating MRI-derived measures with genomic and transcriptomic data may improve biological interpretation by linking imaging signals to molecular pathways. This approach can help identify mechanisms such as altered iron transport and storage, mitochondrial dysfunction, oxidative stress and neuroinflammation. These mechanistic insights may support prioritization of therapeutic targets for excess brain iron in Parkinson’s disease and related neurodegenerative disorders.
Iron-sensitive MRI has moved brain iron research from a largely post-mortem field into an in vivo research domain. Quantitative susceptibility mapping, R2* mapping and susceptibility-weighted imaging can detect regional differences in magnetic susceptibility that are strongly influenced by tissue iron content, particularly in deep grey
matter structures such as the substantia nigra, putamen, caudate and globus pallidus [5]. These methods provide a measurable, spatially resolved phenotype that can be related to ageing, motor function, cognitive decline and neurodegenerative disease risk [6]. However, iron-sensitive MRI remains biologically indirect and non-specific. Increased susceptibility may reflect ferritin-bound storage, neuromelanin-associated iron, mitochondrial iron dysregulation, oligodendrocyte or myelin-related iron, microglial activation, vascular change or tissue injury. As such, while iron-sensitive MRI identifies where susceptibility changes occur, molecular data are needed to help explain why they occur and what they mean.
A central challenge in interpreting nigrostriatal iron deposition is distinguishing physiological ageing from pathological dysregulation. Iron is required for oxidative phosphorylation, neurotransmitter synthesis, myelin maintenance and many enzymatic processes. Regional increases in iron-sensitive MRI signal may therefore reflect normal metabolic demand, adaptive iron storage or age-related changes in tissue composition rather than direct toxicity. Dopaminergic neurons in the substantia nigra have high energetic requirements and intrinsic vulnerability to oxidative stress, rendering them potentially sensitive to disturbances in iron handling. However, elevated iron-sensitive signal does not prove that iron is driving neuronal injury. It may instead reflect compensatory sequestration, altered ferritin expression, microglial responses, neuromelanin-associated binding or downstream effects of degeneration. The question is “when does age-related iron handling become maladaptive?”
Iron biology is therapeutically attractive because it is chemically active, disease-relevant and potentially modifiable. In the ageing nigrostriatal system, iron has plausible links to oxidative stress, mitochondrial dysfunction, dopamine metabolism, lysosomal impairment, neuroinflammation and lipid peroxidation. However, the therapeutic goal is not simply to reduce total brain iron. Iron is biologically necessary, regionally heterogeneous and compartmentalized across distinct cell types and molecular forms. A reduction in iron-sensitive MRI signal does not necessarily mean that the most relevant component of iron biology has been modified, while a therapy that improves iron trafficking, storage, mitochondrial handling or lipid oxidative stress may be biologically meaningful even if total susceptibility changes are modest. The disappointing results of broad-spectrum iron chelation [7] likely reflect both off-target depletion of essential iron pools and substantial patient heterogeneity. These findings underscore the need to determine which molecular forms and cell types carry the pathogenic signal, the optimal timing for intervention and which pathways can be safely and selectively modulated. Future trials should therefore stratify participants using baseline QSM measures combined with polygenic or omics-derived iron vulnerability scores.
Omics approaches could transform nigrostriatal iron from a radiological observation into sets of candidate genes, pathways and therapeutic hypotheses. Genome- and exome-wide association studies of brain iron traits [8, 9] have identified associated loci, but post-GWAS
functional genomics is essential for biological interpretation. Transcriptome-wide association studies (TWAS), splicing analyses, proteome-wide association studies, methylation QTL integration, colocalization and fine-mapping can help identify genes whose genetically regulated expression, splicing, protein abundance or methylation may influence nigrostriatal iron-sensitive phenotypes [10, 11]. Iron biology spans multiple systems: iron transport and storage, ferritinophagy and lysosomal release, mitochondrial iron-sulfur cluster biology, dopamine metabolism, oxidative stress responses, lipid peroxidation and ferroptosis pathways, and neuroinflammation. These distinct signatures suggest different therapeutic strategies: modulating import/export for trafficking defects, targeting NCOA4-ferritinophagy for intracellular iron release, supporting mitochondrial function for energy and iron-sulfur defects, or enhancing antioxidant defense for ferroptosis vulnerability.
Cell-type annotation is critical because the same regional MRI signal may originate from dopaminergic neurons, oligodendrocytes, astrocytes, microglia, endothelial cells or perivascular spaces. A practical prioritization framework should rank candidate targets according to: (1) strength of association with the iron-sensitive imaging phenotype, (2) colocalization with expression, splicing, protein or methylation QTLs, (3) overlap with disease or ageing-related genetic risk, (4) cell-type-specific expression in relevant nigrostriatal populations (leveraging single-nucleus RNA-seq), (5) druggability or pathway tractability, and (6) supportive evidence from post-mortem tissue or experimental models. Key challenges remain, including limited power in current imaging-omics cohorts, incomplete cell-type resolution, and under-representation of non-European ancestries. Multi-ancestry studies and spatial transcriptomics will be essential for robust translation. Nevertheless, nigrostriatal iron accumulation can serve as an entry point for omics-guided therapeutic target discovery.
Nigrostriatal iron deposition is a compelling phenotype with potential for therapeutic intervention. Consistent with recent discussion of resilience-based approaches to ageing, its value may lie in helping identify molecular states that influence vulnerability or resilience in ageing nigrostriatal circuits [12]. Iron-sensitive MRI has made this phenotype measurable in vivo, but measurement alone cannot determine whether iron accumulation reflects physiological adaptation, pathological dysregulation, compensatory sequestration or downstream injury. This distinction is important because iron is both biologically necessary and potentially harmful. Integrating regional iron-sensitive imaging with genomics, transcriptomics, splicing, proteomics, metabolomics and cell-type-resolved data offers a path toward molecular stratification. Such approaches can identify pathways that regulate maladaptive iron handling, distinguish candidate mechanisms from secondary responses and prioritize therapeutic targets.
Declarations
Author contributions
The author contributed solely to the article.
Availability of data and materials
Not applicable.
Financial support and sponsorship
This work was funded by the National Medical Research Council of Singapore (MOH-001378-00 / CNIG23jan-0008).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Use of AI and AI assisted Technologies
No AI tools were used in the preparation of this article.
References
1. Kasar G, Rasal P, Patil C, Mahajan S, & Upaganlawar A. Proteostasis in aging: mechanistic insights and therapeutic opportunities. Aging Pathobiol Ther, 2025, 7(1): 25-43. [Crossref]
2. Rangareddy H, Fryer J, Shetty A, & Rajhi H. Inflammaging and microbiota: the intersection of aging, inflammation, and gut health. Aging Pathobiol Ther, 2025, 7(2): 85-98. [Crossref]
3. Salmon A, editor. Targeting metabolic function to improve aging and healthspan. Abstracts presented at the 2024 Masoro-Barshop conference on aging. October 10–13, 2024; Mayan Dude Ranch in Bandera, TX, USA. Ant Publishing; 2024, 6(4): 141-151. [Crossref]
4. Abolarin P, Olaoluwa P, Ajibare A, Asuku A, & Ayilara G. A mechanistic narrative review of phytochemical enhancement of mitophagy in Parkinson’s disease: from PINK1/Parkin pathways to dopaminergic neuroprotection. Aging Pathobiol Ther, 2026, 8(1): 17-32. [Crossref]
5. Welton T, Hartono S, Shih Y, Schwarz S, Xing Y, Tan E, et al. Ultra-high-field 7T MRI in Parkinson’s disease: ready for clinical use?—a narrative review. Quant Imaging Med Surg, 2023, 13(11): 7607-7620. [Crossref]
6. Hartono S, Chen R, Welton T, Tan A, Lee W, Teh P, et al. Quantitative iron-neuromelanin MRI associates with motor severity in Parkinson’s disease and matches radiological disease classification. Front Aging Neurosci, 2023, 15: 1287917. [Crossref]
7. Devos D, Labreuche J, Rascol O, Corvol J, Duhamel A, Guyon D, et al. Trial of deferiprone in Parkinson’s disease. N Engl J Med, 2022, 387(22): 2045-2055. [Crossref]
8. Wang C, Martins-Bach A, Alfaro-Almagro F, Douaud G, Klein J, Llera A, et al. Phenotypic and genetic associations of quantitative magnetic susceptibility in UK Biobank brain imaging. Nat Neurosci, 2022, 25(6): 818-831. [Crossref]
9. Gong W, Fu Y, Wu BS, Du J, Yang L, Zhang Y, et al. Whole-exome sequencing identifies protein-coding variants associated with brain iron in 29,828 individuals. Nat Commun, 2024, 15(1): 5540-5556. [Crossref]
10. Welton T, Saw W, Zhou Z, Sun Q, Mai A, Teo T, et al. Identification of genes underlying nigrostriatal iron accumulation: transcriptome-wide association study of iron-sensitive brain MRI. EBioMedicine, 2026, 123: 106076. [Crossref]
11. Gu S, Welton T, Sun Q, Wu Y, Tan E, & Zhou Z. The integration of genome-wide and transcriptome-wide association studies in neurodegenerative diseases: opportunities, challenges, and current methodological innovations. Brief Bioinform, 2025, 26(4): bbaf350. [Crossref]
12. Ladiges W. Predicting resilience to aging with physical stressors. Aging Pathobiol Ther, 2025, 7(3): 142-143. [Crossref]