Open Access | Research Article
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Thioredoxin down-regulation in the cytosol in thioredoxin 2 transgenic mice did not have beneficial effects to extend lifespan in male C57BL/6 mice
* Corresponding author: Yuji Ikeno, MD, PhD
Mailing address: Barshop Institute for Longevity and Aging
Studies, The University of Texas Health Science Center at San
Antonio, 4939 Charles Katz Dr, San Antonio, TX 78229, USA.
E-mail: ikeno@uthscsa.edu
Received: 22 November 2020 / Accepted: 20 December 2020
DOI: 10.31491/APT.2020.12.041
Abstract
Background: This study was conducted to test the effects of thioredoxin (Trx)1 down-regulation in Trx2 transgenic [Tg(TXN2)
+/0] mice on lifespan and age-related diseases. Our previous study with Tg(TXN2)
+/0 mice showed
that mitochondrial Trx overexpression produced minimal life-extending effects with a slightly elevated severity
of lymphoma and another study with a limited number of Trx1KO mice showed that there was a slight reduction
of neoplastic lesions. Thus, this study was aimed to test if reduced Trx1 expression in combination with elevated
Trx2 has beneficial effects on lifespan in mice by attenuating age-related diseases, specifically cancer.
Methods: Trx2 hemizygous transgenic and Trx1 heterozygous knockout mice [Tg(TXN2)
+/0 x Trx1KO] were
generated for survival and cross-sectional pathology experiments.
Results: Tg(TXN2)
+/0 x Trx1KO mice showed significantly higher (approximately 1.5- to 3-fold) Trx2 levels and
significantly less (approximately 50% less) Trx1 levels in all of the tissues we examined compared to wildtype (WT) littermates. Trx1 down-regulation along with Trx2 overexpression did not change the levels of
glutathione or other major antioxidant enzymes. Male Tg(TXN2)
+/0 x Trx1KO mice demonstrated only a slight
extension of lifespan in the early part of life and no significant effects on the later part of life were observed,
which was similar to our previous study with Tg(TXN2)
+/0 mice. Tg(TXN2)
+/0 x Trx1KO mice had similar tumor
burden, disease burden, incidence and severity of lymphoma, and severity of glomerulonephritis compared to
WT mice at 22-26 months.
Conclusions: Our findings suggest that the combined Trx down-regulation in cytosol along with upregulation
in mitochondria of Tg(TXN2)
+/0 x Trx1KO mice did not provide beneficial effects on aging, i.e., extend the lifespan or reduce age-related pathology compared to WT mice.
Keywords
Thioredoxin, transgenic mouse, knockout mouse, aging, cancer
Introduction
Possible beneficial roles of thioredoxin (Trx) in aging and
age-related diseases have been investigated in multiple
studies, including several performed in our laboratory
[1-5]. Trx is of interest in aging research because of its
unique ability to alter oxidative stress and redox state,
thereby affecting redox-sensitive signaling and subsequent
effects on pathophysiology. This small protein (12 kDa)
has two redox-active cysteine residues in the active center
(Cys-Gly-Pro-Cys). It plays an important role as the reductant for various molecules via thiol-disulfide exchange
reactions [6-12]. These rapid and readily reversible reactions are ideal to control protein function by changing the
redox state of structural or catalytic SH groups of proteins.
Our laboratory has conducted aging studies with mice
overexpressing or downregulating Trx in the cytosol and/
or mitochondria to test the effects of Trx on aging and
age-related diseases [2-5]. First, we examined the effects
of Trx1 (cytosolic Trx) overexpression on aging with two
lines of transgenic mice [Tg(act-TXN)
+/0 and Tg(TXN)
+/0
mice] [2, 3]. Survival experiments with these mice showed
that overexpression of Trx1 slightly extended the earlier
part of life but did not show significant effects on maximum lifespans. Interestingly, Trx1 overexpression was accompanied by accelerated cancer development in the later
part of life [2, 3].
These initial studies with Trx1 transgenic mice led us to
conduct another survival study with mice overexpressing Trx in mitochondria (Trx2) because the importance of
antioxidant overexpression in mitochondria in aging was
strongly suggested by the study by Schriner et al [13]. The
survival study using Trx2 transgenic mice [Tg(TXN2)
+/0]
showed that Trx overexpression in mitochondria had a
slight but not significant extension (approximately 8-9%)
of mean, median, and 10th percentile lifespans compared
to WT mice. Tg(TXN2)
+/0 mice also showed a slightly
higher severity of lymphoma compared to WT mice [4].
Because we have not observed significant effects on
lifespan, specifically in the latter part of life, by Trx overexpression in cytosol or mitochondria alone, we further
tested if it is necessary to have changes in the levels of
Trx in both the cytosol and mitochondria. One of the potential reasons Trx2 overexpression did not significantly
extend lifespan could be due to its effects on age-related
tumor formation [4]. Both Trx1 and Trx2 transgenic mice
showed a tendency to accelerate tumor development with
age [2-4]. Our previous study with Trx1KO mice showed
that reduced Trx1 did not change the lifespan, i.e., no extension or shortening; however, there was a subtle reduction of age-related cancer based on limited cross-sectional
pathological analyses [5]. These observations led us to the
hypothesis that reducing Trx1 could prevent age-related
tumor development in Tg(TXN2)
+/0 mice, resulting in an
exteded lifespan compared to WT mice.
Thus, the purpose of this study is to examine the effects
of reduced Trx1 along with Trx2 overexpression on aging
and age-related diseases. We conducted a survival study
using mice overexpressing Trx2 and down-regulating
Trx1 [Tg(TXN2)
+/0 x Trx1KO mice]. We report that the
combined Trx down-regulation in cytosol and upregulation in mitochondria did not extend the lifespan, although
these mice did show a slight extension (approximately
30.7%) of lifespan in the early stage of life. Age-related
pathology in Tg(TXN2)
+/0 x Trx1KO mice was similar to
WT mice. Therefore, our results suggest that reduced Trx1
along with Trx2 overexpression did not have significant
effects on lifespan or age-related diseases including tumor
development in male C57BL/6 mice.
Materials and Methods
Animals and animal husbandry
Tg(TXN2) +/0 mice were generated using the human thioredoxin 2 gene [a PAC clone (RP5-1119A7), Children's Hospital Oakland Research Institute's (CHORI) BACPAC Resources Center (BPRC), Oakland, CA] as previously described [4]. Trx1 heterozygous knockout mice were generated with an ES clone purchased from Lexicon Pharmaceuticals, Inc. (OST452454) as previously described [5]. Male heterozygous Trx1KO mice were crossed to female hemizygous Trx2 transgenic mice [Tg(TXN2) +/0] to generate Tg(TXN2) +/0 x Trx1KO and WT control mice. These mice were fed ad libitum with a commercial chow (Teklad Diet LM485: Madison, WI) and free access to acidified (pH = 2.6-2.7) filtered reverse osmosis water. The amount of chow removed from the cage hopper and the spillage (the chow on the bottom of the cage) were weighed monthly to accurately measure the food consumption. All of the mice were weighed monthly. The mice were maintained pathogen-free in microisolator units on Tek FRESH® ultra laboratory bedding. Sentinel mice housed in the same room and exposed weekly to bedding collected from the cages of experimental mice were sacrificed on receipt and every six months thereafter for monitoring of viral antibodies (Mouse Level II Complete Antibody Profile CARB, Ectro, EDIM, GDVII, LCM, M. Ad-FL, M. Ad-K87, MCMV, MHV, M. pul., MPV, MVM, Polyoma, PVM, Reo, Sendai; BioReliance, Rockville, MD). All tests were negative.
Determination of Trx2 expression
The mitochondrial fraction obtained from several tissues (liver, kidney, brain) from young (4-6 months old) Tg(TXN2) +/0 x Trx1KO and WT mice were used to measure thioredoxin 2 (Trx2) levels by Western blot analysis with rabbit anti-Trx2 polyclonal antibody (Catalog No. LF-PA0012; LabFrontier, Seoul, South Korea) [4, 14]. After incubation with the primary antibodies, membranes were incubated with the respective peroxidase-linked secondary antibodies (Catalog No. P0217; Dako, Carpinteria, CA). Chemiluminescence was detected using the ECL Western blot detection kit (Amersham Biosciences Corp., Piscataway, NJ).
Trx1 levels
Trx1 levels were measured using cytosolic fractions obtained from tissues (liver, kidney, brain) of young (4-6 months old) Tg(TXN2) +/0 x Trx1KO and WT mice. Western blot analysis using goat anti-human Trx1 polyclonal antibodies (Catalog No. 705; American Diagnostica, Inc., Greenwich, CT) was performed as previously described [2, 3]. Total Trx1 (both oxidized and reduced forms) levels were measured by these antibodies. After incubation with the primary antibody, membranes were incubated with the peroxidase-linked secondary antibody (Catalog No. P0449; Dako, Carpinteria, CA). Chemiluminescence was detected with an ECL Western blot detection kit (Amersham Biosciences Corp., Piscataway, NJ).
Total glutathione levels
The Bioxytech GSH-420 kit (Catalog No. 21023; Oxis International, Inc., Foster City, CA) was used to measure the levels of total glutathione in several tissues (liver, kidney) from young (4-6 months old) Tg(TXN2) +/0 x Trx1KO and WT mice.
Determination of major antioxidant enzyme activities: Cu/ZnSOD, MnSOD, glutathione peroxidase, and catalase
Major antioxidant enzymes [Cu/ZnSOD, MnSOD, glutathione peroxidase (GPx), and catalase] activities were measured in tissue homogenates obtained from the liver and kidney of young (4-6 months old) Tg(TXN2) +/0 x Trx1KO and WT mice. The antioxidant enzymatic activity assays were conducted with the supernatants. GPx activity in tissue homogenates was measured by the assay as previously described [15]. The Catalase-520TM assay kit (OxisResearchTM, Portland, OR) was used to measure catalase activity by measuring the decomposition of hydrogen peroxide at 520 nm. MnSOD and Cu/ZnSOD levels were determined using activity gels [16, 17]. Images of the gels were analyzed by ImageQuant software.
Survival study
Mice in the survival groups were allowed to live out their lives, and the lifespan for individual mice was determined by recording the age of spontaneous death. A survival study was conducted with 35 Tg(TXN2) +/0 x Trx1KO and 35 WT male mice. The survival curves comparison was statistically analyzed by the log-rank test [18]. The data for mean, 75th percentile, median, and 10th percentile (when 90% of the mice had died) survival were calculated for Tg(TXN2) +/0 x Trx1KO and WT mice. The mean survivals for Tg(TXN2) +/0 x Trx1KO and WT male mice were compared by performing a Student's t-test upon log-transformed survival times. The median and 10th percentile survivals for Tg(TXN2) +/0 x Trx1KO and WT mice were compared using a score test adapted from Wang et al. [19].
Cross-sectional pathological assessment
Tg(TXN2) +/0 x Trx1KO (n = 21) and WT (n = 40) male mice were assigned to the cross-sectional pathological analyses at 22-26 months of age. The gross pathological examinations to detect the visible tumors and other agerelated lesions were conducted with each animal. Then, the following organs and tissues were excised and preserved in 10% buffered formalin: brain, pituitary gland, heart, lung, trachea, thymus, aorta, esophagus, stomach, small intestine, colon, liver, pancreas, spleen, kidneys, urinary bladder, reproductive system (prostate, testes, epididymis, and seminal vesicles), thyroid gland, adrenal glands, parathyroid glands, psoas muscle, knee joint, sternum, and vertebrae. Any other tissues with gross lesions were also excised. After the fixation, tissues were processed conventionally, embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin-eosin. Histological classifications in aging mice were conducted after the diagnosis of each histopathological change was made [20, 21]. A list of pathological lesions for both neoplastic and non-neoplastic diseases was constructed for each mouse. These histopathological data were utilized to determine the tumor burden, disease burden, and severity of each lesion in each mouse [1-4, 21-23].
Statistical analysis
Unless otherwise specified, all experiments were done at least in triplicate. Data were expressed as means ± SEM and were analyzed by the non-parametric test ANOVA. All pair-wise contrasts were computed using Tukey error protection at 95% CI unless otherwise indicated. Differences were considered statistically significant at p < 0.05.
Results
Levels of Trx1 and Trx2 in tissues from Tg(TXN2) +/0 x Trx1KO mice
The levels of Trx1 and Trx2 in tissues (liver, kidney, brain) from young (4-6 months old) Tg(TXN2) +/0 x Trx1KO and WT mice were measured using Western blot analysis. The Trx2 protein levels were significantly higher (approximately 1.3- to 3-fold) in all three tissues examined in young Tg(TXN2) +/0 x Trx1KO mice compared to their WT littermates (Figure 1A; p < 0.05). Levels of Trx1 were significantly lower (approximately 50% less) in all three tissues examined in young Tg(TXN2) +/0 x Trx1KO mice compared to their WT littermates (Figure 1B; p < 0.05).
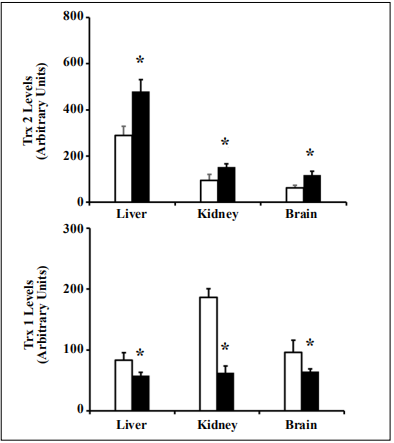
Figure 1. Trx2 and Trx1 levels in young Tg(TXN2) +/0 x Trx1KO mice and their WT littermates. Trx2 and Trx1 protein levels were determined by Western blot in various tissues of 4 to 6 month old Tg(TXN2) +/0 x Trx1KO and WT mice. Trx2 level was significantly (1.5- 3 fold) higher in the tissues (liver, kidney, and brain) of 4 to 6 month old Tg(TXN2) +/0 x Trx1KO (closed bars) and WT mice (open bars) (A, *p < 0.05). Trx1 was significantly lower (approximately 50% less) in the tissues (liver, kidney, and brain) of Tg(TXN2) +/0 x Trx1KO (closed bars) and WT littermates (open bars) (B, *p < 0.05). The data are the mean SEM from three to five mice.
Levels of total glutathione and major antioxidant enzymes in tissues from Tg(TXN2) +/0 x Trx1KO mice
To examine whether the reduced Trx1 along with increased levels of Trx2 affects the levels of glutathione,
which has similar biological functions to Trx, we measured total glutathione levels in liver and kidney from
young (4-6 months old) Tg(TXN2)
+/0 x Trx1KO and WT
mice. The data in Figure 2 show that levels of total glutathione in the tissues (liver, kidney) were similar between
Tg(TXN2)
+/0 x Trx1KO and WT mice at 4-6 months (Figure 2; p > 0.05).
Tg(TXN2)
+/0 x Trx1KO and WT mice showed similar activity/levels for major antioxidant enzymes (Cu/ZnSOD,
MnSOD, GPx, and catalase) in the liver and kidney at 4-6
months (Figure 3; p > 0.05).
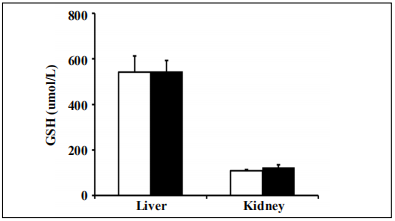
Figure 2. Levels of total glutathione in Tg(TXN2) +/0 x Trx1KO and WT mice. Total glutathione levels were measured in the liver and kidney of young 4 to 6 month Tg(TXN2) +/0 x Trx1KO mice (closed bar) and WT mice (open bar). No significant difference was observed in total glutathione levels in Tg(TXN2) +/0 x Trx1KO compared to WT mice. The data are the mean SEM from three to five mice.
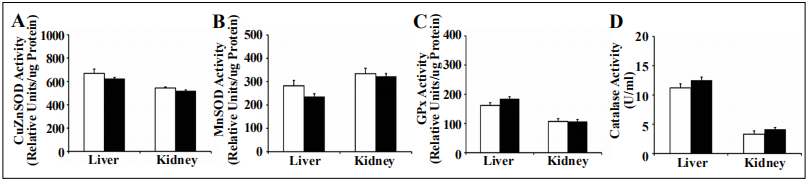
Figure 3. Cu/ZnSOD, MnSOD, GPx, and catalase activity in Tg(TXN2) +/0 x Trx1KO and WT mice. The activities of Cu/ZnSOD (A), MnSOD (B), GPx (C), and catalase (D) were measured in the liver and kidney of 4 to 6 month old Tg(TXN2) +/0 x Trx1KO (closed bar) and WT (open bar) mice. Cu/ ZnSOD, MnSOD, GPx, and catalase activities were similar between Tg(TXN2) +/0 x Trx1KO and WT mice. Data are the mean SEM of three mice.
Survival curves, body and organ weights, and food consumption
The survival study was conducted with male Tg(TXN2) +/0 x Trx1KO (n = 35) and WT (n = 35) mice (Figure 4). The survival curve of Tg(TXN2) +/0 x Trx1KO mice was not significantly different from WT mice (Log-rank: p = 0.45). The mean, 75th percentile, median, and 10th percentile survival for 1) WT mice were 746, 470, 763, and 1,117 days; and 2) Tg(TXN2) +/0 x Trx1KO mice were 835, 678, 791, and 1,141 days, respectively (Figure 4). Tg(TXN2) +/0 x Trx1KO mice had slightly longer mean (10.7), 75th percentile (30.7%), median (3.5%), and 10th percentile (2.1%) lifespans compared to WT mice, although these differences were not statistically significant (p > 0.05). Tg(TXN2) +/0 x Trx1KO mice had similar body and organ weights to WT mice at 4-6 months of age (Table 1). Tg(TXN2) +/0 x Trx1KO and WT mice had similar food intake (data not shown).
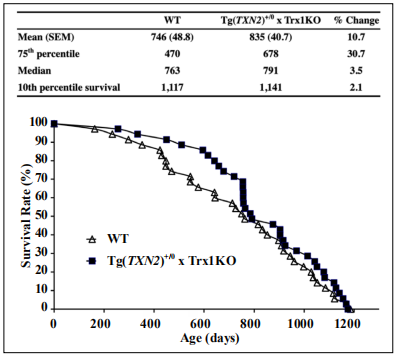
Figure 4. Survival curves of Tg(TXN2) +/0 x Trx1KO and WT mice. The survival curves of Tg(TXN2) +/0 x Trx1KO (closed squares) and WT (open triangles) mice are presented. The cohort consists of 35 Tg(TXN2) +/0 x Trx1KO mice and 35 WT male mice. Although Tg(TXN2) +/0 x Trx1KO mice slightly extended the lifespan in the early stage of life (30.7% extension at 75th percentile), the survival curves were not significantly different between Tg(TXN2) +/0 x Trx1KO and WT mice.
Table 1
Body and Organ Weights of 4-6M Tg(TXN2)
+/0 x Trx1KO mice.
| WT (n=3) | Tg(TXN2) +/0 x Trx1KO (n=4) | |
|---|---|---|
| 29.214±2.14 | 27.279±1.695 | |
| Liver (g) | 1.474±0.069 | 1.447±0.069 |
| Spleen (g) | 0.168±0.012 | 0.165±0.012 |
| Pancreas (g) | 0.287±0.035 | 0.180±0.019 |
| Heart (g) | 0.090±0.017 | 0.085±0.012 |
| Lung (g) | 0.189±0.026 | 0.182±0.006 |
| Left Kidney (g) | 0.229±0.024 | 0.216±0.013 |
| Right Kidney (g) | 0.246±0.024 | 0.245±0.009 |
| Left Testicle (g) | 0.130±0.011 | 0.115±0.008 |
| Right Testicle (g) | 0.127±0.007 | 0.115±0.011 |
| Brain (g) | 0.458±0.015 | 0.455±0.011 |
Cross-sectional pathology
The cross-sectional pathological analyses were conducted
with 21 Tg(TXN2)
+/0 x Trx1KO mice and 40 WT mice
(22-26 months old). The major age-related pathology
was a neoplastic disease, and commonly observed tumors
were lymphoma, hemangioma/hemangiosarcoma (liver
and spleen), adenocarcinoma (lung), and hepatocellular
carcinoma (liver), which is consistent with previous pathology results from the mice that have C57BL/6 genetic
background [1-4, 21].
The tumor burden, which is the total number of different
types of tumors for each mouse, in both Tg(TXN2)
+/0 x
Trx1KO and WT groups is presented in Figure 5A. The
tumor burden of Tg(TXN2)
+/0 x Trx1KO mice was similar
to WT mice (p > 0.05). The incidence of lymphoma was
9% less in Tg(TXN2)
+/0 x Trx1KO (76%) than WT (85%)
mice, and severity of lymphoma was also similar between
Tg(TXN2)
+/0 x Trx1KO (2.50) and WT (2.74) mice (Figure
5B; p > 0.05).
We also compared the severity of glomerulonephritis, a
major non-neoplastic disease, in Tg(TXN2)
+/0 x Trx1KO
and WT mice. The severity of glomerulonephritis was
slightly higher in Tg(TXN2)
+/0 x Trx1KO (1.38) than WT
(1.05) mice, although the difference was not statistically
significant (Figure 5C; p > 0.05).
The disease burden (total number of histopathological
changes in a body) is a good index of tissue and cell injury, which increases with age. Importantly, long-lived mice (e.g., calorie-restricted, Ames, and GHRKO mice)
attenuate its age-related increase [20, 22, 23]. The disease
burden was slightly less in Tg(TXN2)
+/0 x Trx1KO (3.67)
than WT (4.15) mice. However, these differences are not
statistically significant (Figure 5D; p > 0.05).
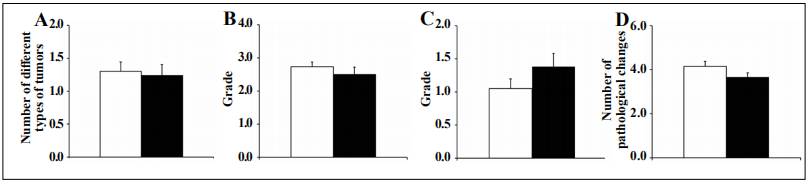
Figure 5. Tumor burden, severity of lymphoma and glomerulonephritis, and disease burden in Tg(TXN2) +/0 x Trx1KO and WT mice. Tumor burden (the number of different types of tumors) (Figure 5A), the severity of lymphoma (Figure 5B), severity of glomerulonephritis (Figure 5C), and disease burden (Figure 5D) in Tg(TXN2) +/0 x Trx1KO (closed bar) and WT (open bar) mice were compared. The tumor burden and severity of lymphoma for the Tg(TXN2) +/0 x Trx1KO mice were similar to WT mice (p > 0.05). Severity of glomerulonephritis was slightly higher in Tg(TXN2) +/0 x Trx1KO than WT mice, however, this difference was not statistically significant (p > 0.05). The disease burden was slightly less in Tg(TXN2) +/0 x Trx1KO (3.67) than WT (4.15) mice, which also did not reach statistical significance.
Discussion
Thioredoxin (Trx) was initially discovered in the early
1960s as the reductant for a variety of enzymes, and many
studies demonstrated its unique roles in biology and physiology [6]. Trx is a hydrogen donor involved in reductive
reactions of enzymes (e.g., ribonucleotide reductase, peroxiredoxin (Prx), and methionine sulfoxide (MetO) [7-12], which play important roles in biology and cellular function. Trx also plays a key role in maintaining a reduced
cellular environment, which provides protection against
oxidative stress and control signaling pathways [24-27].
Trx mainly localizes in the cytosol (hTrx1) [28] and mitochondria (hTrx2) [29] in human cells. Studies with mice
null for either Trx1 or Trx2 showed that these mice are
embryonically lethal, which demonstrated that the presence of Trx1 and Trx2 is essential for mammalian cells/
tissues [30, 31]. Because substantial evidence indicates
Trx and thioredoxin interacting protein (Txnip) regulate
essential cellular functions [6, 32], there is much scientific
interest in how Trx could regulate mammalian aging.
Effects of overexpressed or downregulated Trx in the
cytosol and/or mitochondria on aging and age-related
diseases have been systemically investigated by our laboratory using transgenic and knockout mice for Trx1 or
Trx2 [1-5, 21]. The first Trx study we conducted involved
survival experiments with two lines of Trx1 transgenic
mice [Tg(act-TXN)
+/0 and Tg(TXN)
+/0 mice] [2, 3]. These survival studies demonstrated that overexpression of Trx1
only showed a slight extension in the earlier part of life,
and no significant effects on maximum lifespan were observed, possibly due to accelerated tumor development in
the later part of life [2, 3].
The next study we conducted was a survival experiment
with Trx2 transgenic mice [Tg(TXN2)
+/0]. This study was
conducted because of the strong indication that the protection of the mitochondria from age-related increases in
oxidative stress may delay aging. The survival curve for
Tg(TXN2)
+/0 mice showed a slight (approximately 8-9%)
extension of mean, median, and 10th percentile lifespans
compared to WT mice, although the extension of lifespan
was not statistically significant. Tg(TXN2)
+/0 mice also
showed a trend that the severity of lymphoma was slightly
higher than WT mice [4].
These studies using Trx1 and Trx2 transgenic mice could
indicate that overexpression of Trx in only one compartment of the cell may not have a significant impact on
aging. Our studies with Trx1 or Trx2 knockout (KO)
mice also showed down-regulation of Trx only in one
compartment of the cell showed little impact on aging [5,
14]. Therefore, we decided to test the combined effects
of Trx expression changes in both the cytosol and mitochondria. Among the possible combinations of changes
(either up-regulation or down-regulation) of Trx1 and
Trx2, we chose to down-regulate Trx1 and overexpress
Trx2 in mitochondria for this study because one of the potential reasons Trx2 overexpression did not significantly
extend lifespan could be due to its effects on age-related
tumor formation [4]. Our previous study showed that Trx1
down-regulation (Trx1KO mice) had a subtle reduction of
age-related tumor incidence [5]. Therefore, the purpose of
this study is to test if reduced Trx1 along with Trx2 overexpression could extend the lifespan and attenuate tumor
formation in Tg(TXN2)
+/0 x Trx1KO mice.
To test the effects of reduced Trx1 along with Trx2 overexpression on aging and age-related diseases, we generated Tg(TXN2)
+/0 x Trx1KO mice by crossing male heterozygous Trx1KO mice with female hemizygous Trx2
transgenic mice [Tg(TXN2)
+/0]. These genetic manipulations did not cause changes in food intake, body weight, or
organ weight. Young (4-6 months old) male Tg(TXN2)
+/0
x Trx1KO mice showed significantly higher Trx2 and significantly reduced Trx1 levels in all the tissues examined compared to WT control mice. The Trx2 and Trx1 levels
were 1.5- to 3-fold higher and approximately 50% less in
all of the tissues examined, respectively. The changes in
Trx2 and Trx1 levels in male Tg(TXN2)
+/0 x Trx1KO mice
did not cause changes in total glutathione levels or the
activities of major antioxidant enzymes (Cu/ZnSOD, MnSOD, GPx, and catalase).
Our survival study demonstrated that the survival curve
of male Tg(TXN2)
+/0 x Trx1KO mice was not significantly different from WT control mice. Interestingly, the
Tg(TXN2)
+/0 x Trx1KO mice showed a slight extension of
lifespan in the early stage of life, i.e., 75th percentile lifespan of Tg(TXN2)
+/0 x Trx1KO mice was 30.7% longer than
WT mice although these changes were not statistically
significant. Mean, median, and 10th percentile lifespans of
Tg(TXN2)
+/0 x Trx1KO mice were similar to WT control
mice. The cross-sectional pathology data showed that both
Tg(TXN2)
+/0 x Trx1KO and WT mice had a similar number of total tumors (tumor burden) [Tg(TXN2)
+/0 x Trx1KO
mice: 1.24; and WT control mice: 1.30]. The incidence of
lymphoma was approximately 10% less in Tg(TXN2)
+/0 x
Trx1KO than WT control mice, however, the severity of
lymphoma was similar between Tg(TXN2)
+/0 x Trx1KO
and WT mice. In addition to neoplastic diseases, the total
number of pathological changes (disease burden) was also
similar between Tg(TXN2)
+/0 x Trx1KO and WT control
mice. The severity of glomerulonephritis, which is one of
the major non-neoplastic lesions, was slightly higher in
Tg(TXN2)
+/0 x Trx1KO than WT control mice (not statistically significant). These pathological observations indicate
that down-regulation of Trx1 along with Trx2 overexpression had little effect on age-related pathology, although
the Tg(TXN2)
+/0 x Trx1KO mice slightly extended the
earlier stage of lifespan (75th percentile) compared to WT
control mice.
The outcome of this study clearly showed that Trx1
down-regulation combined with Trx2 overexpression did
not support our working hypothesis, i.e., Tg(TXN2)
+/0 x
Trx1KO mice did not significantly extend lifespan and attenuate age-related diseases. To examine the synergistic
effects of Trx1 and Trx2, we also conducted a survival
study with mice overexpressing Trx1 and Trx2 [1]. This
study demonstrated that overexpression of Trx in both the cytosol and mitochondria shortened the lifespan and
accelerated cancer development in male C57BL/6 mice,
which was contrary to our expectations [1]. Furthermore,
the preliminary data from our current ongoing study show
that reduced thioredoxin levels in both the cytosol and mitochondria [Trx1KO x Trx2KO mice] slightly extend the
lifespan in both male and female C57BL/6 mice, which
is accompanied by suppressed cancer formation [5]. This
study, along with the studies involving either overexpressing or down-regulating Trx1 or Trx2 strongly suggest that
synergetic overexpression or down-regulation of Trx1 and
Trx2 may be required to have significant effects on lifespan and age-related pathology, specifically cancer.
Declarations
Acknowledgments
We acknowledge the Pathology Core in the San Antonio Nathan Shock Center (P30-AG013319) for pathological analyses and funded by the VA Merit Review grant from the Department of Veterans Affairs.
Conflict of interest
The authors declare no conflict of interests.
References
1. Cunningham G M, Flores L C, Roman M G, et al. Thioredoxin overexpression in both the cytosol and mitochondria accelerates age-related disease and shortens lifespan in male C57BL/6 mice. Geroscience, 2018, 40(5-6): 453-468.
2. Flores L C, Roman M G, Cunningham G M, et al. Continuous overexpression of thioredoxin 1 enhances cancer development and does not extend maximum lifespan in male C57BL/6 mice. Pathobiology of Aging & Age-Related Diseases, 2018, 8(1): 1533754.
3. Pérez V I, Cortez L A, Lew C M, et al. Thioredoxin 1 overexpression extends mainly the earlier part of life span in mice. Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences, 2011, 66(12): 1286-1299.
4. Roman M G, Flores L C, Cunningham G M, et al. Thioredoxin overexpression in mitochondria showed minimum effects on aging and age-related diseases in male C57BL/6 mice. Aging Pathobiology and Therapeutics, 2020, 2(1): 20-31.
5. Roman M G, Flores L C, Cunningham G M, et al. Thioredoxin and aging: What have we learned from the survival studies?. Aging Pathobiology and Therapeutics, 2020, 2(3): 126-133.
6. Arnér E S J, Holmgren A. Physiological functions of thioredoxin and thioredoxin reductase. European Journal of Biochemistry, 2000, 267(20): 6102-6109.
7. Brot N, Weissbach L, Werth J, et al. Enzymatic reduction of protein-bound methionine sulfoxide. Proceedings of the National Academy of Sciences, 1981, 78(4): 2155-2158.
8. Brot N, Weissbach H. Peptide methionine sulfoxide reductase: biochemistry and physiological role. Peptide Science, 2000, 55(4): 288-296.
9. Chae H Z, Kang S W, Rhee S G. Isoforms of mammalian peroxiredoxin that reduce peroxides in presence of thioredoxin. Methods in enzymology, 1999, 300: 219-226.
10. Chae H Z, Kim H J, Kang S W, et al. Characterization of three isoforms of mammalian peroxiredoxin that reduce peroxides in the presence of thioredoxin. Diabetes research and clinical practice, 1999, 45(2-3): 101-112.
11. Kim K, Kim I H, Lee K Y, et al. The isolation and purification of a specific" protector" protein which inhibits enzyme inactivation by a thiol/Fe (III)/O2 mixed-function oxidation system. Journal of Biological Chemistry, 1988, 263(10): 4704-4711.
12. Levine R L, Berlett B S, Moskovitz J, et al. Methionine residues may protect proteins from critical oxidative damage. Mechanisms of ageing and development, 1999, 107(3): 323-332.
13. Schriner S E, Linford N J, Martin G M, et al. Extension of murine life span by overexpression of catalase targeted to mitochondria. science, 2005, 308(5730): 1909-1911.
14. Pérez V I, Lew C M, Cortez L A, et al. Thioredoxin 2 haploinsufficiency in mice results in impaired mitochondrial function and increased oxidative stress. Free Radical Biology and Medicine, 2008, 44(5): 882-892.
15. Sun Y, Elwell J H, Oberley L W. A simultaneous visualization of the antioxidant enzymes glutathione peroxidase and catalase on polyacrylamide gels. Free radical research communications, 1988, 5(2): 67-75.
16. Beauchamp C, Fridovich I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Analytical biochemistry, 1971, 44(1): 276-287.
17. Williams M D, Van Remmen H, Conrad C C, et al. Increased oxidative damage is correlated to altered mitochondrial function in heterozygous manganese superoxide dismutase knockout mice. Journal of Biological Chemistry, 1998, 273(43): 28510-28515.
18. Andersen P K, Borgan O, Gill R D, et al. Statistical models based on counting processes. Springer Science & Business Media, 2012.
19. Wang C, Li Q, Redden D T, et al. Statistical methods for testing effects on “maximum lifespan”. Mechanisms of ageing and development, 2004, 125(9): 629-632.
20. Bronson R T, Lipman R D. of Age Related Lesions in Dietary Restricted Laboratory Mice. Growth, Development & Aging, 1991, 55: 169-184.
21. Ikeno Y, Hubbard G B, Lee S, et al. Housing density does not influence the longevity effect of calorie restriction. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 2005, 60(12): 1510-1517.
22. Ikeno Y, Bronson R T, Hubbard G B, et al. Delayed occurrence of fatal neoplastic diseases in Ames dwarf mice: correlation to extended longevity. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 2003, 58(4): B291-B296.
23. Ikeno Y, Hubbard G B, Lee S, et al. Reduced incidence and delayed occurrence of fatal neoplastic diseases in growth hormone receptor/binding protein knockout mice. Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences, 2009, 64(5): 522-529.
24. Abate C, Patel L, Rauscher F J, et al. Redox regulation of fos and jun DNA-binding activity in vitro. Science, 1990, 249(4973): 1157-1161.
25. Galter D, Mihm S, DRÖGE W. Distinct effects of glutathione disulphide on the nuclear transcription factors κB and the activator protein‐1. European Journal of Biochemistry, 1994, 221(2): 639-648.
26. Takagi Y, Mitsui A, Nishiyama A, et al. Overexpression of thioredoxin in transgenic mice attenuates focal ischemic brain damage. Proceedings of the National Academy of Sciences, 1999, 96(7): 4131-4136.
27. Toledano M B, Leonard W J. Modulation of transcription factor NF-kappa B binding activity by oxidationreduction in vitro. Proceedings of the National Academy of Sciences, 1991, 88(10): 4328-4332.
28. Tagaya Y, Maeda Y, Mitsui A, et al. ATL-derived factor (ADF), an IL-2 receptor/Tac inducer homologous to thioredoxin; possible involvement of dithiol-reduction in the IL‐2 receptor induction. The EMBO journal, 1989, 8(3): 757-764.
29. Spyrou G, Enmark E, Miranda-Vizuete A, et al. Cloning and expression of a novel mammalian thioredoxin. Journal of Biological Chemistry, 1997, 272(5): 2936-2941.
30. Matsui M, Oshima M, Oshima H, et al. Early embryonic lethality caused by targeted disruption of the mouse thioredoxin gene. Developmental biology, 1996, 178(1): 179-185.
31. Nonn L, Williams R R, Erickson R P, et al. The absence of mitochondrial thioredoxin 2 causes massive apoptosis, exencephaly, and early embryonic lethality in homozygous mice. Molecular and cellular biology, 2003, 23(3): 916-922.
32. Yoshihara E, Masaki S, Matsuo Y, et al. Thioredoxin/ Txnip: redoxisome, as a redox switch for the pathogenesis of diseases. Frontiers in immunology, 2014, 4: 514.