Open Access | Mini Review
This work is licensed under a Creative
Commons Attribution-ShareAlike 4.0 International License.
Role of fibro-adipogenic progenitors in skeletal muscle aging
* Corresponding author: Xiliang Shang
Mailing address: Department of Sports Medicine, Huashan Hospital, Fudan University, Shanghai 200040, China.
Email: sxl00720@163.com
* Corresponding author: Zhiwen Luo
Mailing address: Department of Sports Medicine, Huashan Hospital, Fudan University, Shanghai 200040, China.
Email: zhiwen.luo_fudan@hotmail.com
This article belongs to the Special Issue: Skeletal Aging; Cellular and Molecular Mechanisms
Received: 23 Feburary 2023 / Revised: 03 May 2023 / Accepted: 10 May 2023 / Published: 28 June 2023
DOI: 10.31491/APT.2023.06.116
Abstract
Maintaining muscle mass is of paramount importance from a clinical perspective as it supports the flexibility, strength, and essential daily tasks that the body requires. Furthermore, muscle plays a role in regulating the body’s metabolic system. Unfortunately, aging can lead to a loss of muscle mass, which can reduce personal independence and quality of life while increasing the risk of developing disease. Fibro-adipogenic progenitor cells (FAPs) are muscle-resident progenitor cells that are essential for maintaining skeletal muscle fiber size and muscle regeneration. These vital FAP functions are mediated by a complex secretome that interacts in a paracrine manner to promote the division and differentiation of muscle satellite cells. Dysregulated differentiation of FAPs can lead to fibrosis, fatty infiltration, muscle atrophy, and poor muscle regeneration. In this article, we review what is currently known about how FAPs function in aging muscle and how they may prevent the onset of muscle wasting and degeneration. Finally, we discuss how FAPs represent a population of cells that can be used as therapeutic targets to improve the health of skeletal and muscle tissues as they age.
Keywords
Aging, fibro-adipogenic progenitors, skeletal muscle, muscle regeneration
Introduction
Aging is now a major concern for the global population,
and the rate of aging is accelerating [1]. One of the hallmarks of aging is a progressive decline in skeletal muscle
mass and strength, leading to an increased incidence of
injury, deconditioning, and even loss of independence and
quality of life [2]. In addition, several studies have found
that decreased muscle regeneration, increased fibrosis,
and fatty infiltration are also associated with aging [3-6].
Aging also leads to an imbalance in muscle homeostasis,
and skeletal muscle homeostasis is maintained by a balance of physical and functional interactions of different
cell types in the muscle niche [7-10]. Indeed, multiple cell
types are involved in maintaining skeletal muscle mass
and homeostasis, including fibro-adipogenic progenitor
cells (FAPs), tenocytes, endothelial cells, smooth muscle
cells, immune cells (B cells, T cells, macrophages, neutrophils), neural or glial cells [11, 12]. When skeletal muscle
homeostasis is perturbed by various pathological factors,
the muscle environment also triggers dynamic changes in
the composition of cell types and functional interactions
between these cells [7, 13, 14]. For these reasons, there is
great interest in understanding the regulation and mechanisms of muscle degeneration in order to develop effective
therapeutic strategies.
Over the past decade, FAPs have been recognized as important regulators of muscle homeostasis and regeneration in healthy muscle, but also in acutely injured skeletal
muscle and pathologically degenerated muscle. FAPs were
first identified in 2010 as muscle-resident progenitor cells
that express PDGFR and primarily give rise to myofibroblasts and adipocytes [15, 16]. Under normal conditions of
muscle regeneration, activated FAPs eventually undergo
apoptosis through mechanisms dependent on macrophagesecreted tumor necrosis factor [17]. However, if apoptosis
does not occur in a timely manner, FAPs can differentiate
into pro-fibrotic fibroblasts and white adipose tissue in
the presence of prolonged inflammatory signals in injured
muscle [14, 18, 19]. Furthermore, crosstalk between FAPs
and other cells in the muscle stem cell (MuSC) ecotone
plays a critical role in restoring and maintaining muscle
structure and function [20-23]. Due to the importance of
FAPs in the regenerative and degenerative muscle environment, balancing FAP activity is essential to promote
effective muscle regeneration without inducing chronic
muscle degeneration.
Here, we review the current understanding of the role of
FAPs in muscle aging and the characterization of FAPs in
aging muscle. We also discuss the plasticity and behavior
of FAPs in the tissue microenvironment. Finally, we highlight the therapeutic potential of FAPs in regenerating aging muscle.
Contribution and mechanism of FAPs in aging
Aging is characterized by a decline in several physiological functions. The regenerative potential of muscle decreases with age, and the progressive loss of skeletal muscle mass is also known as sarcopenia [24]. Age-related sarcopenia is an important health problem that is closely associated with impaired muscle regeneration, impaired adaptive response to exercise training, and disorders of muscle metabolic regulation [25]. Meanwhile, degeneration and atrophy of aging muscles are associated with increased fibrosis, fatty infiltration, and low-grade chronic inflammation [6]. In human and mouse muscles, FAPs are thought to be the cellular origin of fibrosis and adipogenesis, leading to chronic inflammation and muscle loss [26, 27]. Liu et al. found significant co-localization of FAPs with adipocyte markers using PDGFRα-GFP reporter mice [19]. Jensen et al. also found similar co-localization when differentiating FAPs into adipocytes and fibroblasts in vitro [21]. These studies are consistent in strongly suggesting that FAPs are an important mediator of adipose tissue infiltration and fibrosis in muscle. The exact mechanisms that allow FAPs to gravitate toward lipogenesis and fibrogenesis are currently unknown, but may include alterations in local signaling, gene expression, and stem cell epigenetics, as well as the presence of baseline differences in subpopulations of FAPs. For example, Moratal et al. found that aging leads to changes in the niche to which FAPs are exposed, creating a more favorable environment for fibrotic or adipogenic differentiation of FAPs [8].
Fibrillation
One of the hallmarks of aging muscle is increased fibrotic tissue. Several studies have shown that as muscle ages, the activity of FAPs is impaired and the number of FAPs and their ability to proliferate decreases, while the tendency for fibrotic differentiation increases [28, 29]. Mueller et al. found that aging induces FAPs to enter a fibrotic state [30], and several intrinsic cellular defects have been shown to contribute to the impaired activity of aging FAPs. A reduction in the truncated variant of the PDGFRα, which acts as a decoy receptor to inhibit the PDGF signaling pathway, has been observed in aged FAPs [29]. In addition, the environment of aging stem cells is known to be more inflammatory than that of young cells [31]. Inflammatory factors such as elevated levels of IL-6, IL-8, IL- 1β, TNF-α, and NF-κβ are known to characterize the aging stem cell environment [32]. These cytokines have been shown to have a significant effect on fibrosis in FAPs (Figure 1). For example, the presence of higher levels of the pro-fibrotic factor TGF-β during the aging process [33], and the TGF-β signaling pathway, a known stimulator of fibrosis in FAPs, is upregulated in injured muscle, with macrophages identified as the major source of TGF-β [17, 34, 35]. In rotator cuff injuries, aging is associated with an increase in fibrosis [36]. An increase in fibrosis is also due to an increase in myostatin levels [37]. Dong et al. have shown that myostatin causes increased proliferation and fibrotic differentiation of FAPs through upregulation of PSmad2/Smad3 [38].
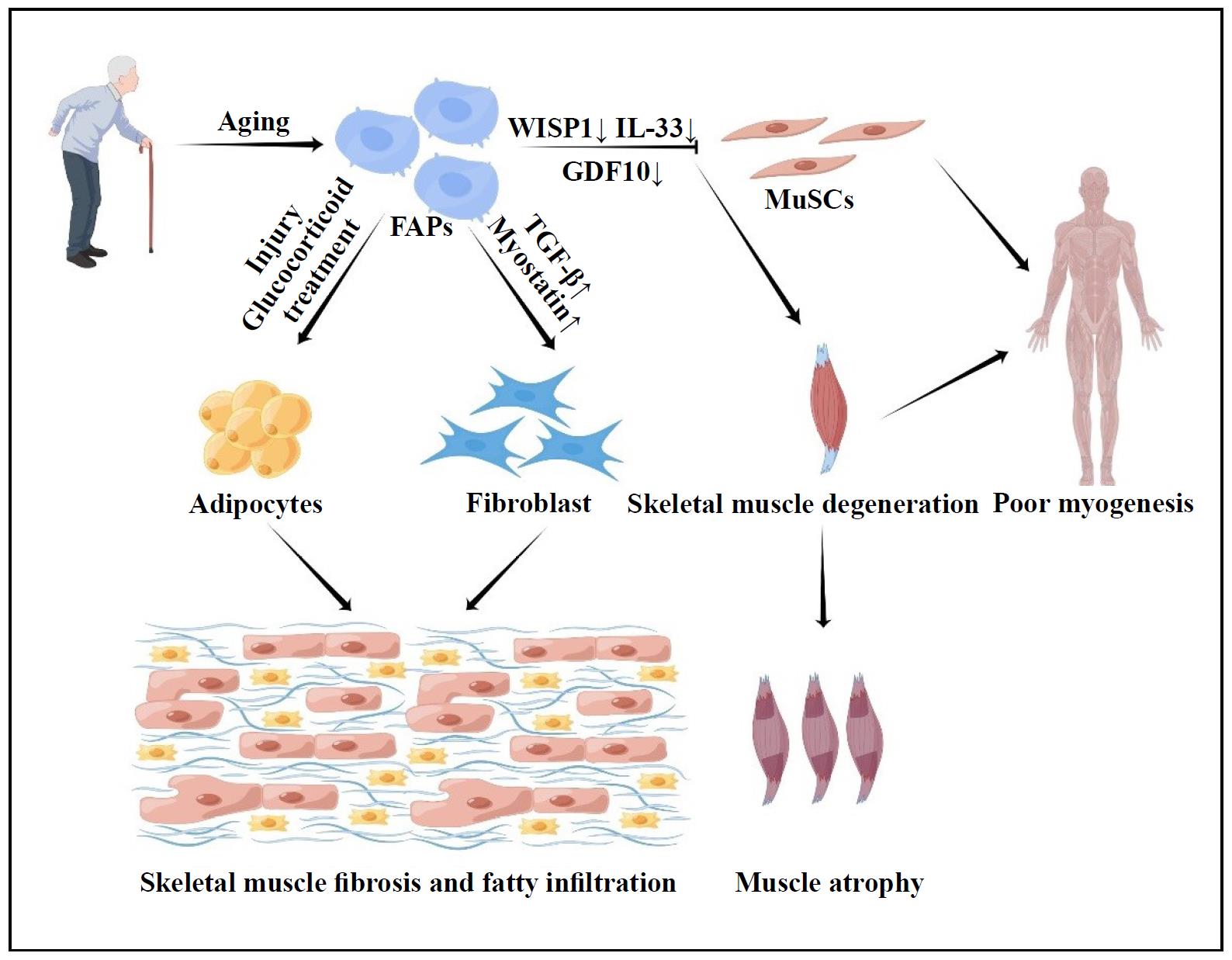
Figure 1. Contribution and mechanism of FAPs in aging. When activated by multiple circumstances, aging FAPs are more prone to fibrosis and adipogenesis. Furthermore, aging FAPs impair MuSc function, resulting in muscle atrophy.
Adipogenic differentiation
As skeletal muscle atrophies, the amount of fat in the muscle increases, a process called myosteatosis. This process is another hallmark of muscle aging. Because of their adipogenic potential, FAPs play a central role in myosteatosis. Adipogenic differentiation pathways in FAPs are stimulated by both injury and glucocorticoid treatment. For example, Itoigawa et al. found increased levels of the adipogenic markers PPARγ and CEBPα in a rat model of rotator cuff tear [39]. The correlation between increased number of FAPs and fatty infiltration with larger tear size suggests that different tear conditions may induce epigenetic changes in FAPs, thereby altering their proliferation and differentiation behavior [40]. Furthermore, a study found that conditioned medium from myogenic progenitors isolated from young individuals increased FAP proliferation and inhibited adipogenic differentiation, whereas conditioned medium from myogenic progenitors isolated from aged donors did not improve FAP proliferation and prevented adipogenic differentiation [8].
Comorbidities
The effect of aging on FAPs is also associated with the presence of comorbidities. For example, the incidence of type 2 diabetes increases dramatically with age [41]. The study by Mogi et al. showed that ectopic fat deposition in regenerated muscle of diabetic mice was derived from FAPs [42]. Insulin resistance in type 2 diabetes leads to overproduction of this hypoglycemic hormone, which is a known inducer of adipogenic differentiation of FAPs in vitro [22]. A recent study found that enhanced conversion of a subset of FAPs to CD90+ FAPs is related to degenerative remodeling of the extracellular matrix in the skeletal muscle of type 2 diabetic patients. CD90+ FAPs exhibit a PDGF-mimetic phenotype with significant clonogenicity, proliferative activity, and extracellular matrix synthesis [26]. Obesity is also common in the elderly. Obesity severely impairs muscle contractility and leads to progressive expansion of adipose tissue and collagen deposition during high fat intake, which may be due to increased number and proliferation of FAPs in chronically obese patients [43].
Secretion factors
Lukjanenko et al. showed that aging impairs the function
of mouse FAPs [29]. Notably, they describe the inability
of aging FAPs to support MuSCs due to reduced secretion of the stromal cell protein WNT1-inducible signaling pathway protein 1 (WISP1) [29]. WISP1 plays an
important role in the asymmetric division of muscle stem
cells and muscle regeneration. Cellular transplantation
of young FAPs into old mice restores the commitment of
muscle stem cells to myogenesis, supporting the role of
FAPs in the dysfunction of myogenesis during aging [29]
(Figure 1). Meanwhile, lower levels of GDF10 are also
expressed in aging FAPs. Uezumi et al. found that in vitro
addition of conditioned medium from transgenic FAPs
overexpressing GDF10 induced myotubular hypertrophy
to a greater extent than conditioned medium from wildtype or GDF10 knockout FAPs, and that administration
of GDF10 to aged mice reversed muscle mass loss and
myofiber atrophy [28] (Figure 1). Muscle regeneration is
also hampered by the ineffective production of paracrine
substances by FAPs. FAPs are the major source of IL-33,
a cytokine associated with type 2 immunity, in monocytes;
however, we produce less of this cytokine as we age, resulting in less Treg accumulation and suboptimal muscle
regeneration [41] (Figure 1).
These findings strongly suggest that age-related changes
affect the ability of FAPs to maintain homeostasis. To
prevent muscle aging and sarcopenia, modulation of FAPderived cues holds great therapeutic promise.
Potential therapeutic role of FAPs in aging
FAP activity and number
Regulation of FAP activity and number may have an impact on how much fibrosis and adipose infiltration occurs after muscle aging because FAPs have the ability to develop into fibroblasts and adipocytes. Numerous studies have actually tested this theory. Lemos et al. discovered that blocking TNF signaling stopped FAP apoptosis, resulting in twice the amount of FAPs and twice the amount of fibrosis after muscle injury. They also showed that nilotinib, a tyrosine kinase inhibitor that targets the TGF signaling pathway, increased FAP apoptosis and decreased fibrosis [17]. Imatinib, a related small molecule inhibitor that inhibits PDGFRα signaling and has been shown in studies to increase grip strength in mice with dystrophic limbs, significantly reduced muscle fibrosis [34]. In the rotator cuff muscle, the small molecule inhibitor CWHM- 12 has been shown to dramatically reduce FAP-induced fibrosis in in vitro assays [21]. Although it appears that reducing the overall amount and activity of FAPs may reduce their downstream pathologies, pharmacological removal of FAPs from muscle tissue runs the risk of muting any beneficial effects they may have (Figure 2). The earliest benefits of FAPs have been demonstrated in muscle regeneration [16, 32]. However, the detrimental effects of their later development into fibroblasts and adipocytes may counteract these early beneficial effects. This would indicate that a better strategy than simply eliminating them would be to alter their behavior to achieve a more benificial phenotype. This would preserve and enhance their original benificial roles.
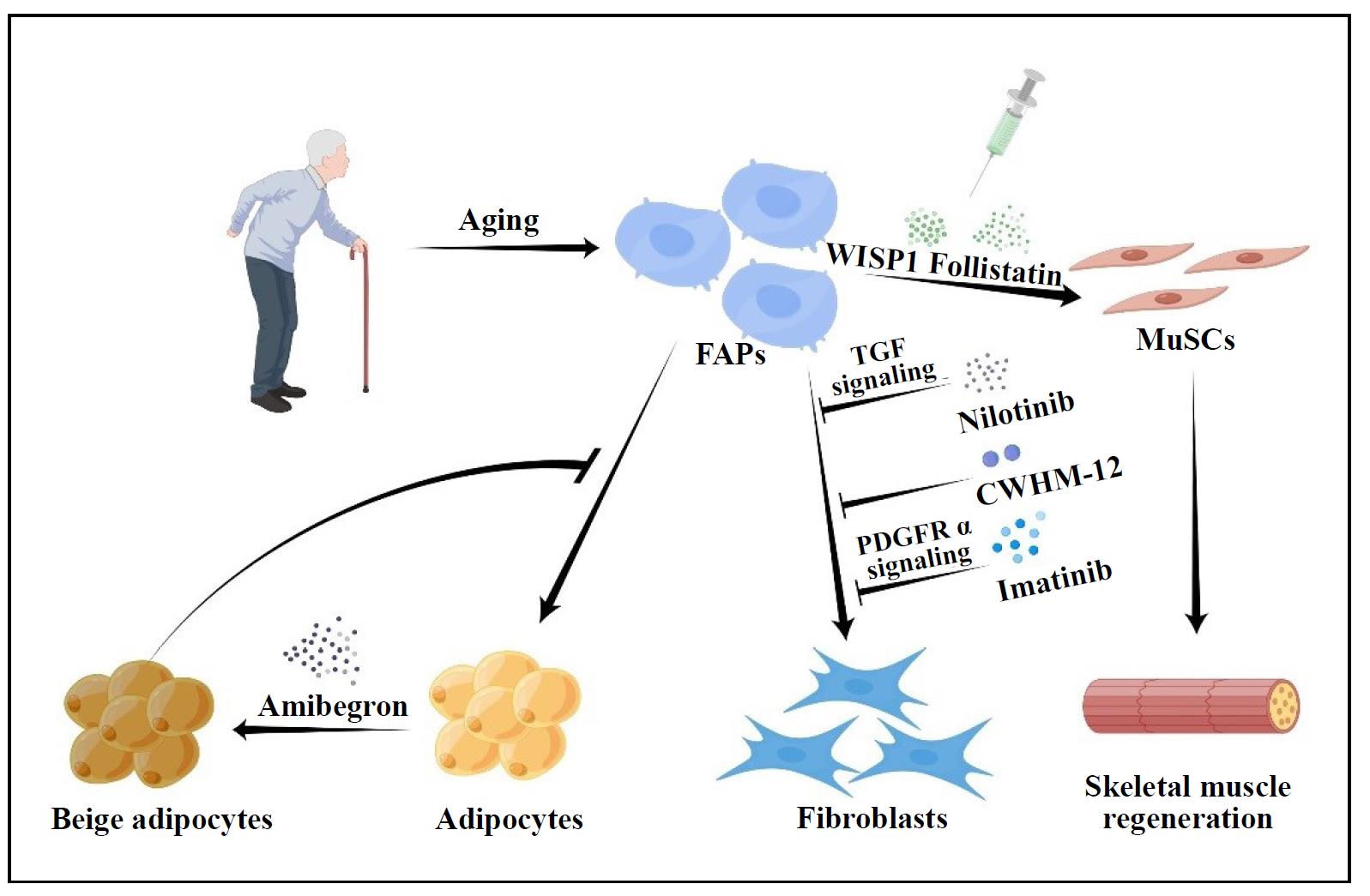
Figure 2. Potential therapeutic role of FAPs in aging. We can play a therapeutic function by decreasing FAPs activity, decreasing FAPs number, synergizing the interaction between FAPs and MuScs, and controlling FAPs differentiation.
Secreted factors of FAPs
Lukjanenko et al. found that in vivo treatment with recombinant WISP1 improved muscle architecture and myofiber cross-sectional area, increased the proportion of newly formed myofibers, and enhanced the early proliferation of aged MuSCs and the commitment of Pax7+ /MyoD+ MuSCs in aged muscle subjected to acute injury [29]. As a result, systemic treatment of WISP1 improves the diminished regenerative capacity of aging muscle, indicating that FAP-secreted molecules may be used as potential therapies to enhance the endogenous capacity of muscle regeneration (Figure 2). Mozzetta et al. demonstrated that aging inhibits the FAP-stimulated production of MuSCderived multinucleated myotubes through co-culture experiments between MuSCs and FAPs. Co-transplantation of FAPs increased MuSC engraftment and muscle regeneration in aged mice, which was also confirmed in vivo [45]. The authors hypothesized that follistatin from FAPs mediates their pro-myogenic effects on MuSCs, which are strengthened by histone deacetylase inhibitor (HDACi) therapy [45] (Figure 2). The production and transport of miRNA-carrying extracellular vesicles by FAPs may be a mechanism by which HDACi-mediated pro-myogenic effects enhance MuSCs [46].
Differentiation tendency of FAPs
Recent research has also focused on the ability of FAPs
to differentiate into a more benificial adipose phenotype
to investigate how to promote muscle regeneration while
reducing adipose infiltration and fibrosis. Through the
production of uncoupled protein 1 (UCP-1), which prevents cellular respiration, brown fat can generate heat. In
this respect, beige fat is similar to brown fat and can also
express UCP-1, but it comes from the same place as white
adipose tissue [47, 48]. There is increasing evidence that
the more metabolically active brown and beige adipose
tissues have a pro-myogenic role in addition to their basic
thermogenic activity [32, 49, 50]. Meyer et al. showed
that beige fat between rotator cuff muscles increases myotube formation in co-culture experiments with myogenic
progenitor cells. Based on this, successful efforts have
been made, such as those by Lee et al., to promote differentiation of FAPs to the beige adipose phenotype. The
prevalence of fibrosis, fat infiltration, atrophy, and gait
impairment in mice with supraspinatus muscle deterioration was significantly reduced by transplantion of beige
adipose differentiated from FAPs derived from UCP-1
reporter mice [51, 52]. Additional in vitro studies have
recently shown that Amibegron-treated human rotator
cuff FAPs tend to differentiate toward a beige adipose
phenotype [40]. Future studies could focus on transplatation methods and pharmaceutical strategies to enhance the
conversion of rotator cuff FAPs to a beige fat phenotype.
These studies could clarify whether the approaches under
this hypothesis reduce fat infiltration and fibrosis in the
muscle and promote myokine secretion to aid in muscle
regeneration (Figure 2).
Collectively, age-related intrinsic and extrinsic changes
affect the regulation of FAPs, which in turn promotes the
formation of fibrotic tissue and hinders muscle regeneration. Further research is needed to evaluate the potential
of therapeutic molecules targeting FAPs to revitalize aged
skeletal muscle.
Conclusion and future directions
Understanding the mechanisms underlying tissue regeneration and degeneration caused by cellular aging is
critical to the maintenance of skeletal muscle health. The
multiple functions of skeletal muscle cell aging in muscle
regeneration and degeneration are increasingly supported
by research. In this review, we discuss the fundamental
relationships between FAPs and aging, and how FAPs influence age-related muscle degeneration and regeneration.
The regulatory function of FAPs in maintaining muscle
growth and function serves as an example of recent developments in our understanding of these proteins. The current data clearly demonstrate that FAPs play a critical role
in the regulation of skeletal muscle homeostasis. Their
ability to differentiate directly determines whether the effects on muscle synthesis and regeneration are positive or
unfavorable, and these effects are carefully controlled by
signaling molecules in the muscle stem cell ecotone.
Skeletal muscle research will evolve with a better understanding of the relationship between FAPs and the pathophysiology and physiology of muscle aging. However,
there are still several issues that need to be addressed.
First, the phenotypes and roles of FAPs that are spatiotemporally modulated during skeletal muscle aging and
degeneration are still unclear; second, what is the role
of FAPs in controlling the initiation and development of
muscle senescence degeneration; Third, how FAPs and
muscle stem cells interact to affect muscle and tissue integrity, and how immune cells and FAPs interact to maintain homeostasis; To reduce or prevent fibrosis, fat infiltration, and degenerative aging of muscle, it may be possible
to use muscle-resident FAPs or their subpopulations as
therapeutic targets. By addressing these issues, therapeutic
strategies targeting FAPs cells in aging skeletal muscle
will be developed.
In summary, there is no question that understanding how
to improve recovery from skeletal muscle damage and
minimize muscle loss in the elderly is critical. Moreover,
while FAPs cells remain largely unknown, a fuller knowledge of the intricate role of muscle aging with FAPs is
essential for the creation of truly effective targeted therapeutics for aging.
Declarations
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 81972125 and 82172510).
Authors’ contributions
Zhiwen Luo and Xiliang Shang contributed to the conception and design of the study. Renwen Wan, Hanli Zhang, and Shan Liu designed and wrote the whole manuscript. Chunmeng Jiao and Hui Chen completed subsequent revisions of the manuscript. Renwen Wan, Hanli Zhang, Shan Liu and Hui Chen collected the references and prepared figures of the manuscript. All authors contributed to manuscript revision, and approved the final version of manuscript.
Conflicts of interest
The authors have declared that no conflicts interest exists.
Availability of data and materials
Not applicable.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
References
1. Hsu W, & Yang FC. Factors Associated with Home Health Aides’ Turnover Intention and Organizational Citizenship Behavior in Long-Term Care Services. Healthcare (Basel), 2022, 10(9): 1743-1753. [Crossref]
2. Bao Z, Cui C, Chow SK, Qin L, Wong RMY, & Cheung WH. AChRs Degeneration at NMJ in Aging-Associated Sarcopenia-A Systematic Review. Front Aging Neurosci, 2020, 12: 597811. [Crossref]
3. Spehar K, Pan A, & Beerman I. Restoring aged stem cell functionality: Current progress and future directions. Stem Cells, 2020, 38(9): 1060-1077. [Crossref]
4. Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, et al. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science, 2007, 317(5839): 807-810. [Crossref]
5. Etienne J, Liu C, Skinner CM, Conboy MJ, & Conboy IM. Skeletal muscle as an experimental model of choice to study tissue aging and rejuvenation. Skelet Muscle, 2020, 10(1): 4-14. [Crossref]
6. Rahman FA, Angus SA, Stokes K, Karpowicz P, & Krause MP. Impaired ECM Remodeling and Macrophage Activity Define Necrosis and Regeneration Following Damage in Aged Skeletal Muscle. Int J Mol Sci, 2020, 21(13): 4575- 4585. [Crossref]
7. Cordani N, Pisa V, Pozzi L, Sciorati C, & Clementi E. Nitric oxide controls fat deposition in dystrophic skeletal muscle by regulating fibro-adipogenic precursor differentiation. Stem Cells, 2014, 32(4): 874-885. [Crossref]
8. Moratal C, Arrighi N, Dechesne CA, & Dani C. Control of Muscle Fibro-Adipogenic Progenitors by Myogenic Lineage is Altered in Aging and Duchenne Muscular Dystrophy. Cell Physiol Biochem, 2019, 53(6): 1029-1045. [Crossref]
9. Wang Y, Yang Z, Yang L, Zou Q, Zhao S, Hu N, et al. Liuweidihuang Pill Alleviates Inflammation of the Testis via AMPK/SIRT1/NF-kappaB Pathway in Aging Rats. Evid Based Complement Alternat Med, 2020, 2020: 2792738. [Crossref]
10. Cutler AA, Dammer EB, Doung DM, Seyfried NT, Corbett AH, & Pavlath GK. Biochemical isolation of myonuclei employed to define changes to the myonuclear proteome that occur with aging. Aging Cell, 2017, 16(4): 738-749. [Crossref]
11. Mashinchian O, Pisconti A, Le Moal E, & Bentzinger CF. The Muscle Stem Cell Niche in Health and Disease. Curr Top Dev Biol, 2018, 126: 23-65. [Crossref]
12. Wosczyna MN, & Rando TA. A Muscle Stem Cell Support Group: Coordinated Cellular Responses in Muscle Regeneration. Dev Cell, 2018, 46(2): 135-143. [Crossref]
13. Chung SW, Kim JY, Yoon JP, Suh DW, Yeo WJ, & Lee YS. Atrogin1-induced loss of aquaporin 4 in myocytes leads to skeletal muscle atrophy. Sci Rep, 2020, 10(1): 14189. [Crossref]
14. Farup J, Madaro L, Puri PL, & Mikkelsen UR. Interactions between muscle stem cells, mesenchymal-derived cells and immune cells in muscle homeostasis, regeneration and disease. Cell Death Dis, 2015, 6(7): e1830. [Crossref]
15. Uezumi A, Fukada S, Yamamoto N, Ikemoto-Uezumi M, Nakatani M, Morita M, et al. Identification and characterization of PDGFRalpha+ mesenchymal progenitors in human skeletal muscle. Cell Death Dis, 2014, 5(4): e1186. [Crossref]
16. Uezumi A, Fukada S, Yamamoto N, Takeda S, & Tsuchida K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nat Cell Biol, 2010, 12(2): 143-152. [Crossref]
17. Lemos DR, Babaeijandaghi F, Low M, Chang CK, Lee ST, Fiore D, et al. Nilotinib reduces muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of fibro/adipogenic progenitors. Nat Med, 2015, 21(7): 786-794. [Crossref]
18. Heredia JE, Mukundan L, Chen FM, Mueller AA, Deo RC, Locksley RM, et al. Type 2 innate signals stimulate fibro/ adipogenic progenitors to facilitate muscle regeneration. Cell, 2013, 153(2): 376-388. [Crossref]
19. Liu X, Ning AY, Chang NC, Kim H, Nissenson R, Wang L, et al. Investigating the cellular origin of rotator cuff muscle fatty infiltration and fibrosis after injury. Muscles Ligaments Tendons J, 2016, 6(1): 6-15. [Crossref]
20. Biferali B, Proietti D, Mozzetta C, & Madaro L. FibroAdipogenic Progenitors Cross-Talk in Skeletal Muscle: The Social Network. Front Physiol, 2019, 10: 1074-1084. [Crossref]
21. Jensen AR, Kelley BV, Mosich GM, Ariniello A, Eliasberg CD, Vu B, et al. Neer Award 2018: Platelet-derived growth factor receptor alpha co-expression typifies a subset of platelet-derived growth factor receptor betapositive progenitor cells that contribute to fatty degeneration and fibrosis of the murine rotator cuff. J Shoulder Elbow Surg, 2018, 27(7): 1149-1161. [Crossref]
22. Uezumi A, Ito T, Morikawa D, Shimizu N, Yoneda T, Segawa M, et al. Fibrosis and adipogenesis originate from a common mesenchymal progenitor in skeletal muscle. J Cell Sci, 2011, 124(Pt 21): 3654-3664. [Crossref]
23. Wang Z, Liu X, Davies MR, Horne D, Kim H, & Feeley BT. A Mouse Model of Delayed Rotator Cuff Repair Results in Persistent Muscle Atrophy and Fatty Infiltration. Am J Sports Med, 2018, 46(12): 2981-2989. [Crossref]
24. Joanisse S, Nederveen JP, Snijders T, McKay BR, & Parise G. Skeletal Muscle Regeneration, Repair and Remodelling in Aging: The Importance of Muscle Stem Cells and Vascularization. Gerontology, 2017, 63(1): 91-100. [Crossref]
25. Burton LA, & Sumukadas D. Optimal management of sarcopenia. Clin Interv Aging, 2010, 5: 217-228. [Crossref]
26. Farup J, Just J, de Paoli F, Lin L, Jensen JB, Billeskov T, et al. Human skeletal muscle CD90(+) fibro-adipogenic progenitors are associated with muscle degeneration in type 2 diabetic patients. Cell Metab, 2021, 33(11): 2201- 2214 e2211. [Crossref]
27. Giuliani G, Vumbaca S, Fuoco C, Gargioli C, Giorda E, Massacci G, et al. SCA-1 micro-heterogeneity in the fate decision of dystrophic fibro/adipogenic progenitors. Cell Death Dis, 2021, 12(1): 122-132. [Crossref]
28. Uezumi A, Ikemoto-Uezumi M, Zhou H, Kurosawa T, Yoshimoto Y, Nakatani M, et al. Mesenchymal Bmp3b expression maintains skeletal muscle integrity and decreases in age-related sarcopenia. J Clin Invest, 2021, 131(1): e139617. [Crossref]
29. Lukjanenko L, Karaz S, Stuelsatz P, Gurriaran-Rodriguez U, Michaud J, Dammone G, et al. Aging Disrupts Muscle Stem Cell Function by Impairing Matricellular WISP1 Secretion from Fibro-Adipogenic Progenitors. Cell Stem Cell, 2019, 24(3): 433-446.e437. [Crossref]
30. Mueller AA, van Velthoven CT, Fukumoto KD, Cheung TH, & Rando TA. Intronic polyadenylation of PDGFRalpha in resident stem cells attenuates muscle fibrosis. Nature, 2016, 540(7632): 276-279. [Crossref]
31. Nilsson MI, Bourgeois JM, Nederveen JP, Leite MR, Hettinga BP, Bujak AL, et al. Lifelong aerobic exercise protects against inflammaging and cancer. PLoS One, 2019, 14(1): e0210863. [Crossref]
32. Joe AW, Yi L, Natarajan A, Le Grand F, So L, Wang J, et al. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat Cell Biol, 2010, 12(2): 153-163. [Crossref]
33. Forsey RJ, Thompson JM, Ernerudh J, Hurst TL, Strindhall J, Johansson B, et al. Plasma cytokine profiles in elderly humans. Mech Ageing Dev, 2003, 124(4): 487-493. [Crossref]
34. Huang P, Zhao XS, Fields M, Ransohoff RM, & Zhou L. Imatinib attenuates skeletal muscle dystrophy in mdx mice. FASEB J, 2009, 23(8): 2539-2548. [Crossref]
35. Davies MR, Lee L, Feeley BT, Kim HT, & Liu X. Lysophosphatidic acid-induced RhoA signaling and prolonged macrophage infiltration worsens fibrosis and fatty infiltration following rotator cuff tears. J Orthop Res, 2017, 35(7): 1539-1547. [Crossref]
36. Sharma AK, Levian B, Shah P, Mosich GM, Husman R, Ariniello A, et al. Aged Mice Demonstrate Greater Muscle Degeneration of Chronically Injured Rotator Cuff. J Orthop Res, 2020, 38(2): 320-328. [Crossref]
37. Bo Li Z, Zhang J, & Wagner KR. Inhibition of myostatin reverses muscle fibrosis through apoptosis. J Cell Sci, 2012, 125(Pt 17): 3957-3965. [Crossref]
38. Dong J, Dong Y, Chen Z, Mitch WE, & Zhang L. The pathway to muscle fibrosis depends on myostatin stimulating the differentiation of fibro/adipogenic progenitor cells in chronic kidney disease. Kidney Int, 2017, 91(1): 119- 128. [Crossref]
39. Itoigawa Y, Kishimoto KN, Sano H, Kaneko K, & Itoi E. Molecular mechanism of fatty degeneration in rotator cuff muscle with tendon rupture. J Orthop Res, 2011, 29(6): 861-866. [Crossref]
40. Feeley BT, Liu M, Ma CB, Agha O, Aung M, Lee C, et al. Human Rotator Cuff Tears Have an Endogenous, Inducible Stem Cell Source Capable of Improving Muscle Quality and Function After Rotator Cuff Repair. Am J Sports Med, 2020, 48(11): 2660-2668. [Crossref]
41. Bellary S, Kyrou I, Brown JE, & Bailey CJ. Type 2 diabetes mellitus in older adults: clinical considerations and management. Nat Rev Endocrinol, 2021, 17(9): 534-548. [Crossref]
42. Mogi M, Kohara K, Nakaoka H, Kan-No H, Tsukuda K, Wang XL, et al. Diabetic mice exhibited a peculiar alteration in body composition with exaggerated ectopic fat deposition after muscle injury due to anomalous cell differentiation. J Cachexia Sarcopenia Muscle, 2016, 7(2): 213-224. [Crossref]
43. Buras ED, Converso-Baran K, Davis CS, Akama T, Hikage F, Michele DE, et al. Fibro-Adipogenic Remodeling of the Diaphragm in Obesity-Associated Respiratory Dysfunction. Diabetes, 2019, 68(1): 45-56. [Crossref]
44. Kuswanto W, Burzyn D, Panduro M, Wang KK, Jang YC, Wagers AJ, et al. Poor Repair of Skeletal Muscle in Aging Mice Reflects a Defect in Local, Interleukin-33-Dependent Accumulation of Regulatory T Cells. Immunity, 2016, 44(2): 355-367. [Crossref]
45. Mozzetta C, Consalvi S, Saccone V, Tierney M, Diamantini A, Mitchell KJ, et al. Fibroadipogenic progenitors mediate the ability of HDAC inhibitors to promote regeneration in dystrophic muscles of young, but not old Mdx mice. EMBO Mol Med, 2013, 5(4): 626-639. [Crossref]
46. Sandonà M, Consalvi S, Tucciarone L, De Bardi M, Scimeca M, Angelini DF, et al. HDAC inhibitors tune miRNAs in extracellular vesicles of dystrophic muscle-resident mesenchymal cells. EMBO Rep, 2020, 21(9): e50863. [Crossref]
47. Bartesaghi S, Hallen S, Huang L, Svensson PA, Momo RA, Wallin S, et al. Thermogenic activity of UCP1 in human white fat-derived beige adipocytes. Mol Endocrinol, 2015, 29(1): 130-139. [Crossref]
48. Sidossis L, & Kajimura S. Brown and beige fat in humans: thermogenic adipocytes that control energy and glucose homeostasis. J Clin Invest, 2015, 125(2): 478-486. [Crossref]
49. Wang Z, Feeley BT, Kim HT, & Liu X. Reversal of Fatty Infiltration After Suprascapular Nerve Compression Release Is Dependent on UCP1 Expression in Mice. Clin Orthop Relat Res, 2018, 476(8): 1665-1679. [Crossref]
50. Gorski T, Mathes S, & Krutzfeldt J. Uncoupling protein 1 expression in adipocytes derived from skeletal muscle fibro/adipogenic progenitors is under genetic and hormonal control. J Cachexia Sarcopenia Muscle, 2018, 9(2): 384-399. [Crossref]
51. Lee C, Liu M, Agha O, Kim HT, Feeley BT, & Liu X. Beige FAPs Transplantation Improves Muscle Quality and Shoulder Function After Massive Rotator Cuff Tears. J Orthop Res, 2020, 38(5): 1159-1166. [Crossref]
52. Lee C, Liu M, Agha O, Kim HT, Liu X, & Feeley BT. Beige fibro-adipogenic progenitor transplantation reduces muscle degeneration and improves function in a mouse model of delayed repair of rotator cuff tears. J Shoulder Elbow Surg, 2020, 29(4): 719-727. [Crossref]