Open Access | Research
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Exploring the complex interplay between anxiety, aging, and behavior in CB6F1 and C57BL/6 mice: implications for cognitive function
# These authors contributed equally.
* Corresponding author: Warren Ladiges
Mailing address: Department of Comparative Medicine, School of Medicine, University of Washington, Seattle, WA, USA.
Email: wladiges@uw.edu
Received: 23 January 2024 / Revised: 19 February 2024 / Accepted: 06 March 2024 / Published: 28 March 2024
DOI: 10.31491/APT.2024.03.133
Abstract
Anxiety is a pervasive emotional response that can profoundly impact well-being and cognitive function in both humans and animals. The relationship between anxiety and aging remains complex and multifaceted. To explore this relationship in more detail, an open-field photobeam system was used to quantify anxietyrelated behaviors in aging CB6F1 and C57BL/6 male mice and to determine associations with aging phenotypes, including short- and long-term memory, grip strength, rotarod performance, and self-motivated wheel running. Results indicated a heightened anxiety in novel environments with increasing age as evidenced by a preference for peripheral areas during the open-field test. Elevated anxiety levels were not associated with decreased cognitive performance, suggesting that anxiety and cognition operate somewhat independently of each other. A negative correlation was observed between anxiety levels and distance run in the voluntary wheel running assessment, while no associations were seen with grip strength or rotarod performance. These observations contribute to a better understanding of anxiety and its consequences in aging mice, providing insights into potential therapeutic interventions aimed at delaying aging through anxiety management.
Keywords
Anxiety, aging, behavioral assessment, cognition, CB6F1 mice, C57BL/6 mice
Introduction
Anxiety is a pervasive emotion that can manifest as an unpleasant state of internal confusion and a sense of fear toward
impending events [1-3]. While anxiety is a common
physiological response to a stressful situation, the constant presence is a sign of a clinical diagnosis of an anxiety disorder
[4-7]. The numerous types of anxiety disorders
share common risk factors but have distinct symptoms, with generalized anxiety disorder (GAD) and obsessivecompulsive disorder (OCD)
being the most common [8, 9]. There is published
evidence of an association between anxiety and memory, with the presence of anxiety serving as a strong predictor of future
cognitive deterioration [10-13]. Symptoms of
memory impairment often emerge along with those of anxiety [14-17].
Anxiety in mice parallels the physiological responses observed in humans, which include increased alertness, avoidance of certain
environments or stimuli, changes in motor activity, changes in social interactions, and physiological changes such as increased
heart rate or blood pressure [18]. Several behavioral tests have been designed to measure
anxiety in mice based on the introduction of novel environments that may elicit anxiety secondary to an intrinsic fear response.
The open-field test (OFT) is a popular paradigm that employs a floor grid and video tracking system to record movements within
predefined grid zones over various time intervals [19]. The photobeam-based variant of
this test allows for increased tracking frequencies, enabling more precise observations of anxiety-related behaviors
[20]. Anxious tendencies in mice are often characterized by an aversion to open and
exposed spaces, leading to increased time spent in the peripheral areas of the testing area while avoiding exploration of the
central region [21, 22].
This study examines the relationship between anxiety and established aging parameters in mice to help understand the associations
and mechanisms underlying the aging process and its effect on various physiological and behavioral aspects. The findings develop
further insight into age-related disorders and contribute to the development of novel therapeutic approaches and interventions to
promote healthy aging.
Materials and methods
Animals
CB6F1 (C57BL/6 × Balb/c F1 cross) and C57BL/6 male mice in age groups of 4, 12, 20, 28 months were obtained from the National Institute on Aging Aged Rodent Colony under contract from by Charles River, Inc. Mice were housed in a specific pathogen-free mouse facility at the University of Washington (UW) main campus in Seattle, WA. The status of the room was monitored under the guidance of the Rodent Health Monitoring Program within the purview of the UW Department of Comparative Medicine. Mice were group housed, up to five per cage, and given nestlets (Ancare Corp, Bellmore, NY, USA) for environmental enrichment. Mice were acclimated for two weeks before starting test procedures. All procedures were approved by the University of Washington IACUC (Animal Care and Use Committee).
Anxiety assessment
An open-field photobeam testing system (OFT) (Columbus Instruments, Inc.) was employed to evaluate anxiety-related behaviors of mice in a novel environment as previously described [23]. The apparatus simulates a standard mouse cage, featuring a clear rectangular container and infrared beams arranged in a grid pattern, three horizontally and four vertically. The open-field photobeam system was configured with two sets of infrared beams to measure both lateral and vertical activity. Beam breaks, which occurred when mice crossed an infrared beam, were counted for each activity. The collected data were subsequently categorized into two distinct zones: the central and peripheral areas of the container. This categorization allowed for the assessment of anxiety levels based on the preference for exploring specific regions. Increased time spent in the central area suggested reduced anxiety, whereas a preference for peripheral regions suggested heightened anxiety. Each mouse was placed inside the testing container for a period of five minutes. This standard duration ensured consistent evaluation of anxiety-related behaviors and minimized potential habituation effects or stress-related responses.
Memory assessment
A radial water tread maze was used to assess short-term and long-term memory [24]. The maze consisted of a circular basin with nine holes, eight decoys leading to dead ends, and one escape hole leading to a dark safety box equipped with a heating pad to simulate a standard mouse cage. The basin contained approximately one inch of water and an overhead light was placed above the cage as an escape incentive. Mice had three trials in the maze for four consecutive days of training, followed by testing on the fifth day and retesting on the twelfth day.
Grip strength assessment
Forelimb paw strength was measured using a grip strength meter [25]. Each mouse was positioned horizontally with its forepaws on a metal grip bar (Columbus Instruments, Inc.), and the mouse was pulled back at a uniform rate until releasing the bar. The machine recorded the maximum force exerted by the mouse for a total of five trials. Mice were weighed on the test day and peak force was expressed relative to body weight to normalize grip strength measurements.
Agility assessment
Agility was assessed using a Rotamax 4/8 rotating bar machine (Columbus Instruments, Inc.) [25]. The machine tested the ability of mice to maintain walking speed on a rotating bar. The initial speed was set at 0 RPM and gradually increased by 0.1 RPM/s over a 5-minute duration until all mice fell off and were detected by a sensor. Time in seconds was recorded for each mouse over three trials.
Voluntary wheel running assessment
Total distance run over three days was measured with a running wheel added to a standard mouse cage [26]. Mice were individually housed in standard cages with a slanted running wheel wirelessly connected to a computer (Med Associates, Inc.). There was a two-day acclimation period with the wheel locked, and on the third day the wheel was unlocked and data collection began. Running distances were continuously monitored over a 72-hour period, with total distances run in kilometers recorded every minute.
Data analysis
All data were grouped according to strain and age. A Shapiro-Wilk test was used to assess data under each group to determine whether there was a normal distribution. For normally distributed data, statistical comparisons were performed using parametric tests. Non-normally distributed data were analyzed using non-parametric tests. The Wilcoxon rank sum test was used to determine significant differences between two groups when the data were not normally distributed. The Kruskal-Wallis H test was used to determine significant differences between multiple groups when the data were not normally distributed. In cases where there was no interaction between factors, but one factor was found to be statistically significant, a posthoc analysis of means was conducted using the Bonferroni adjustment method. Spearman's rank test was used to measure the strength of association between aging and each parameter, while the point-biserial correlation coefficient was used for assessing the association between a continuous variable and a binary variable. All statistical analyses were performed using GraphPad Prism (version 10.0.3).
Results
Age-related differences in total movement in the open field test were detected in CB6F1 mice
The results of the two-way ANOVA on total movement in the open field test (OFT) revealed significant main effects of both strain (F (1,84) = 4.313, P < 0.0001) and age (F (1,84) = 29.19, P < 0.05) with no significant interaction between strain and age (F (1,84) = 1.678, P = 0.199). Post-hoc analysis indicated that older mice at 20 to 28 months of age exhibited prolonged periods of movement compared to younger mice at 4 to 12 months of age in CB6F1 but not C57BL/6 (B6) strains (Figure 1A & B, respectively). Furthermore, older CB6F1 mice displayed more extensive movement compared to their B6 cohorts (Figure 1C).
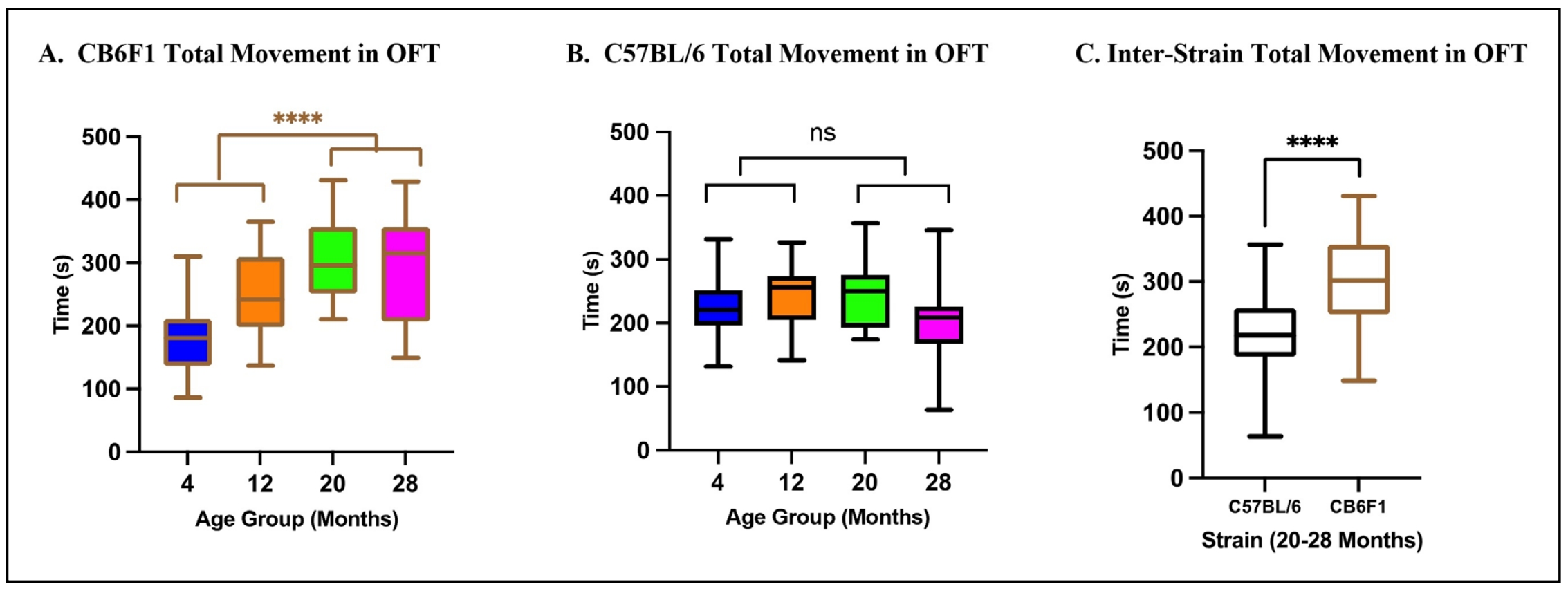
Figure 1. Total movement in the OFT. (A) CB6F1 mice at 20 and 28 months of age exhibited significant differences when compared to 4 and 12-month-old cohorts and showed a positive association between movement and age. (B) B6 mice at the same age groups did not show the same significant differences and age-related increase in movement as seen in CB6F1 mice. (C) Comparison of older B6 and CB6F1 mice showed a significant difference in total movement (****P < 0.0001, ns = not significant (P > 0.05), N = 19-28/cohort, OFT = open-field photobeam testing system).
Mice showed area preferences in the open field test in an age and strain-dependent manner
Older CB6F1 mice exhibited significantly more time in both central and peripheral areas than their younger counterparts as
revealed by the results of a one-way ANOVA with significant main effects of both central (F (3,82) = 8.144, P < 0.0001)
and peripheral (F (3,82) = 10.30, P < 0.0001) area preferences with respect to age after post-hoc analyses
(Figure 2A & B, respectively). Older B6 mice
displayed such distinctions for central areas, as demonstrated by the results of a one-way ANOVA (F (3,77) = 7.587, P <
0.0005), but not for peripheral areas (F (3,77) = 1.526, P = 0.214) after post-hoc analyses
(Figure 2C & D, respectively).
CB6F1 mice at 28 months of age allocated significantly more time in peripheral areas than 28-month-old B6 mice
(Figure 2E). A two-way ANOVA of time spent in the peripheral area revealed a significant
interaction between strain and age (F (3,159) = 5.292, P < 0.005) with posthoc analysis. These observations
collectively suggest that with increasing age, CB6F1 mice have a higher likelihood than B6 mice for prolonged occupancy in
peripheral areas.
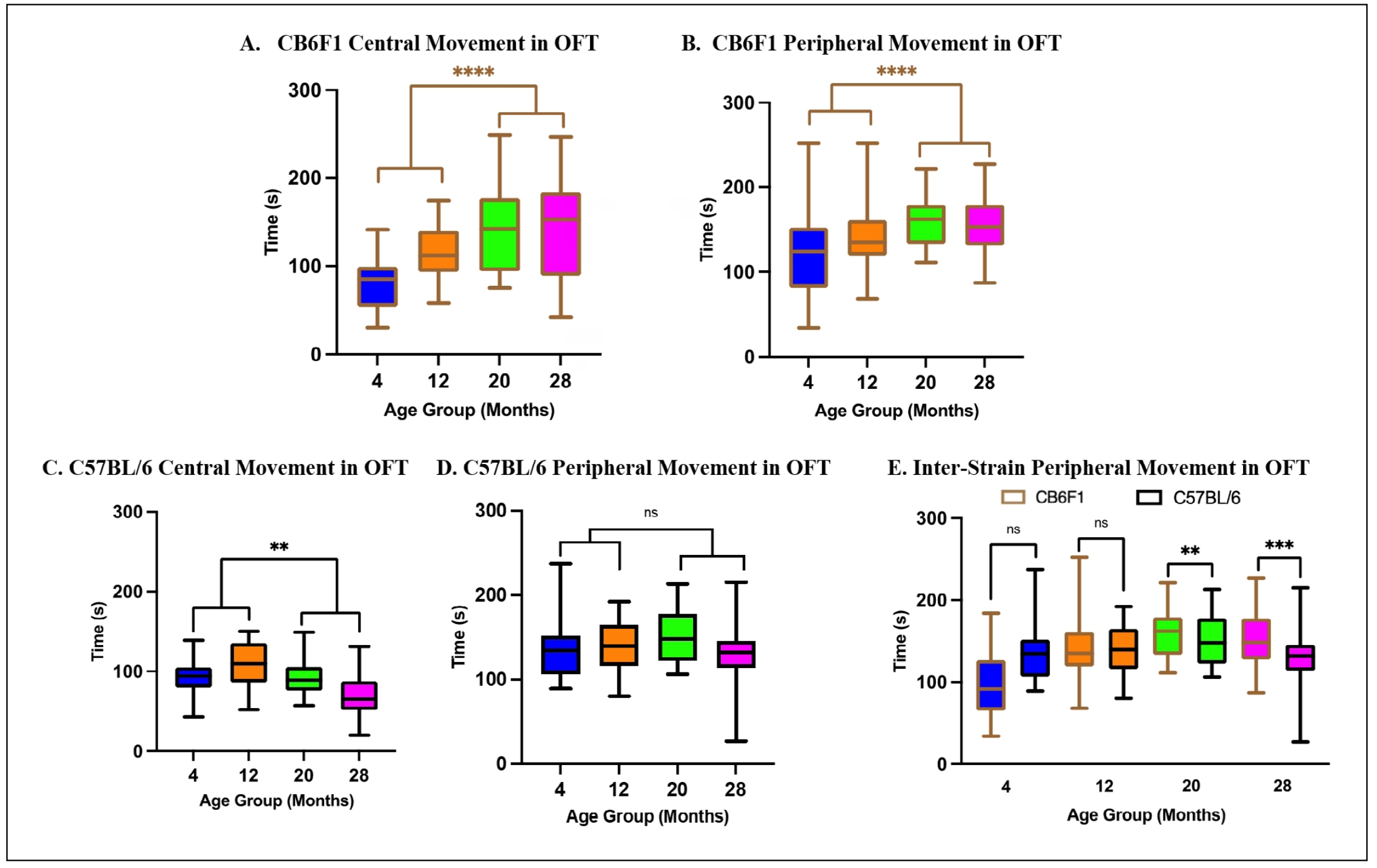
Figure 2. Total time spent in central and peripheral areas. (A-B) Older CB6F1 mice spent significantly more time in central and peripheral areas than their younger counterparts. (C-D) Older B6 mice spent significantly more time in central but not in peripheral areas compared to their younger counterparts. (E) CB6F1 mice at 20- and 28-months of age spent significantly more time in peripheral areas compared to B6 mice (**P < 0.01, ***P < 0.001, ****P < 0.0001, ns = not significant (P > 0.05), N = 19-28/cohort, OFT = open-field photobeam testing system).
Time spent rearing was age dependent in CB6F1 mice
An interaction between time spent rearing and age was demonstrated by a Kruskal-Wallis test for both CB6F1 (H = 19.69, df = 3, P < 0.0005) and B6 mice (H = 9.355, df = 3,P < 0.05), although post-hoc analysis indicated that older CB6F1 mice spent significantly more time rearing than their younger counterparts (Figure 3A), while time spent rearing was not age dependent in B6 mice (Figure 3B). Time spent rearing was area dependent as both CB6F1 and C57BL/6 mice spent significantly more time rearing in the periphery compared to the central regions as demonstrated by Kruskal-Wallis tests ([H = 83.83, df = 7, P < 0.0001], [H = 136.3, df = 7, P < 0.0001], respectively) with post-hoc analysis (Figure 3C & D, respectively). A two-way ANOVA revealed significant main effects of age (F (1,159) = 20.64, P < 0.0001) but not strain (F (3,159) = 1.368, P = 0.255) or interaction between strain and age (F (3,159) = 1.452, P = 0.230) on preference for rearing over activity. Overall, both CB6F1 and B6 mice spent significantly less time rearing than moving laterally (Figure 3E & F, respectively).
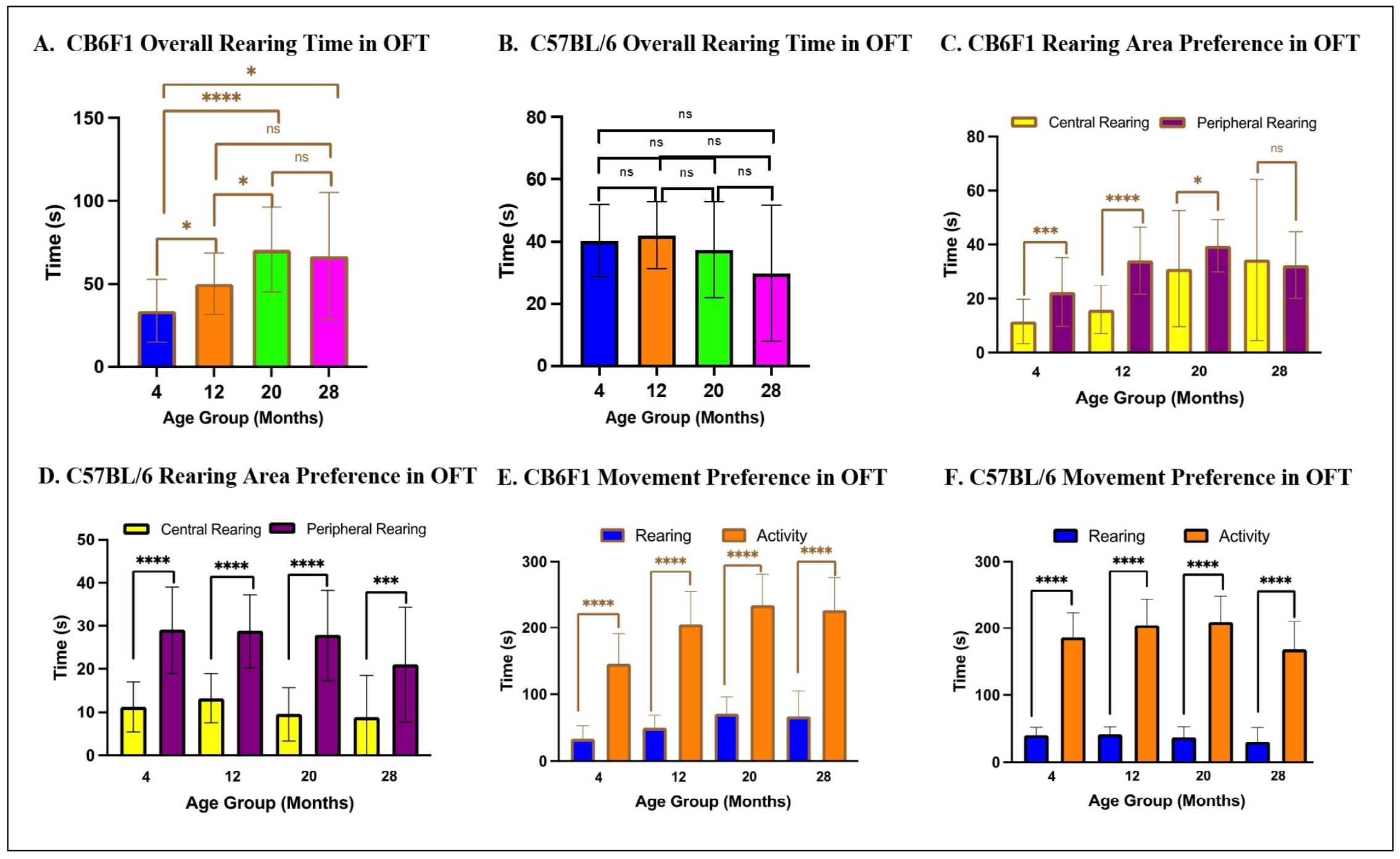
Figure 3. Time spent in each area analyzed according to age and strain. (A-B) Older CB6F1 mice tended to spend more time rearing compared to younger counterparts, while no age-dependent preference was observed in C57BL/6 mice. (C-D) Rearing was area-dependent, as both CB6F1 and C57BL/6 mice tended to rear in peripheral areas for longer compared to central areas. (E-F) CB6F1 and C57BL/6 across all age groups preferred activity compared to rearing (*P < 0.05, ***P < 0.001, ****P < 0.0001, ns = not significant (P > 0.05), N = 19-28/cohort, OFT = open field photobeam testing system).
Memory was associated with age but not peripheral area preference in CB6F1 mice
Multiple linear regression analysis revealed that age was a significant predictor of long-term memory (ß= 3.02, P <
0.0005), with post-hoc analysis indicating that older CB6F1 mice exhibited a significant increase in escape times on day 12 of the
radial water tread maze (Figure 4A). However, the open field ratio (OFR) was not a
significant predictor of long-term memory for any age group (ß= -53.58, P = 0.466)
(Figure 4C-F), and the overall model did not
account for a significant portion of the variance in long-term memory scores (R2 = 0.162, F (2,75) = 7.234, P =
0.093). OFR was calculated as time spent in peripheral areas divided by total recorded time. Long-term memory was quantified by the
time spent to find the escape on day 12.
Regarding short-term memory, the multiple linear regression analysis revealed that age was a significant predictor (ß=2.98,
P < 0.0001), with post-hoc analysis indicating that older CB6F1 mice exhibited a significant increase in escape times
on day 5 of the radial water tread maze (Figure 4B). However, OFR was not a significant
predictor of short-term memory for any age group (ß= -99.18, P = 0.110)
(Figure 4C-F). The overall model accounted
for a significant proportion of the variance in short-term memory scores (R2 = 0.208, F (2,82) = 10.78, P <
0.0001). Short-term memory was measured by the time spent to find the escape on day 5.
For C57BL/6 mice, multiple linear regression and posthoc analyses revealed that neither age (ß = 0.895,P = 0.281)
(Figure 5A) nor OFR (ß = 180.9, P = 0.067)
(Figure 5C-F) were significant predictors of
long-term memory, with the overall model accounting for a significant portion of the variance in long-term memory scores
(R2 = 0.081, F (2,75) = 3.291, P < 0.05). However, for short-term memory, age was a significant predictor
(ß = 2.029, P < 0.05) (Figure 5B), but not OFR (ß = 127.0, P = 0.144)
(Figure 5C-F), with the overall model accounting
for a significant portion of the variance in short-term memory scores (R2 = 0.151, F (2,78) = 6.911, P <
0.005). Post-hoc analysis indicated that older C57BL/6 mice exhibited a significant increase in escape times on day 5 of the radial
water tread maze (Figure 5B). Overall, there were no statistically significant
correlations between memory performance and peripheral area preference across age groups in C57BL/6 mice.
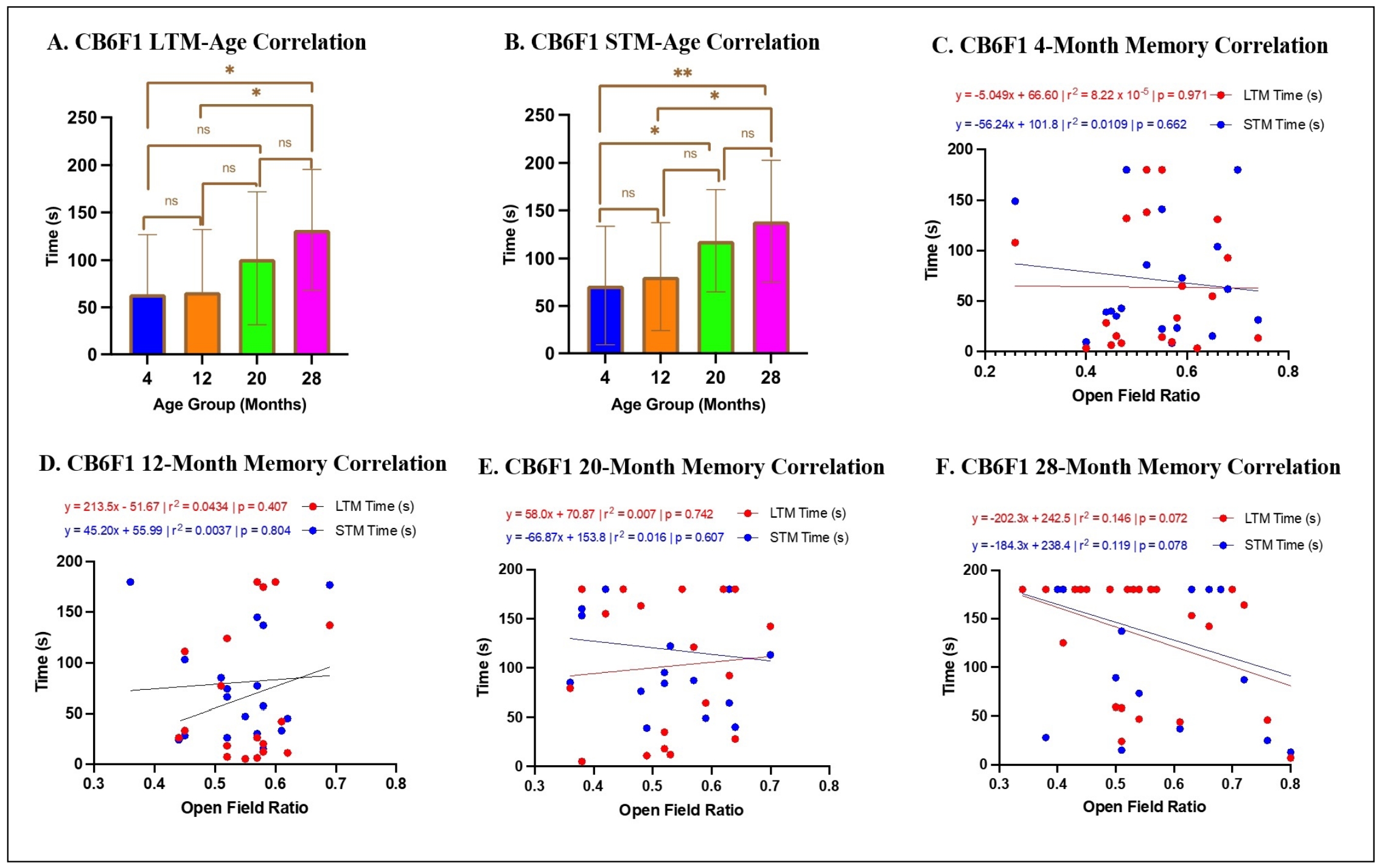
Figure 4. Relationship between memory and OFR in CB6F1 mice. (A-B) There were statistically significant differences across CB6F1 age groups for both long- and short-term memory. (C-F) There were no significant correlations between either long- or short-term memory with anxiety scores (*P < 0.05, **P < 0.01, ns = not significant (P > 0.05), N = 19-28/cohort, LTM = long-term memory, STM = short-term memory, OFR = open field ratio).
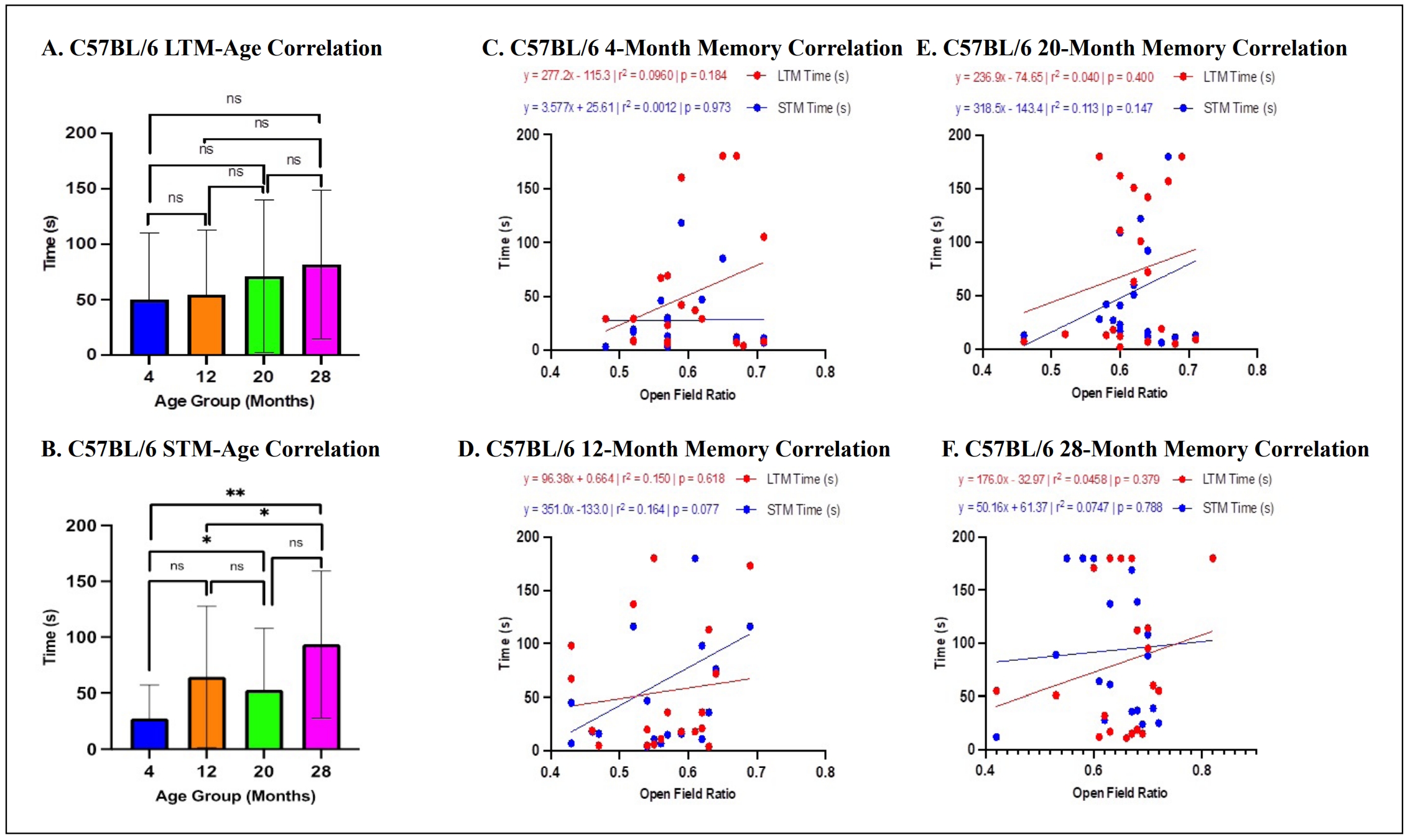
Figure 5. Relationship between memory and OFR in C57BL/6 mice. (A-B) There were statistically significant differences across C57BL/6 age groups for short- but not long-term memory. (C-F) There were no significant correlations between either long- or short-term memory with anxiety scores (*P < 0.05, **P < 0.01, ns = not significant (P > 0.05), N = 19-28/cohort, LTM = long-term memory, STM = short-term memory, OFR = open field ratio).
Peripheral area preference was associated with reduced running distance in 28-month-old C57BL/6 mice, but no associations were found with grip strength or rotarod in either strain
For CB6F1 mice in voluntary wheel running, the multiple linear regression analysis revealed that age was a significant predictor of distance traveled (ß= -0.501, P < 0.0001), with post-hoc analyses indicating that older CB6F1 mice exhibited a significant reduction in self-motivated running distance (Figure 6A). However, OFR was not a significant predictor of distance traveled (ß= -3.661, P = 0.659) (Figure 6B-E). The overall model accounted for a significant portion of the variance in distance traveled (R2 = 0.332, F (2,65) = 16.12, P < 0.0001).
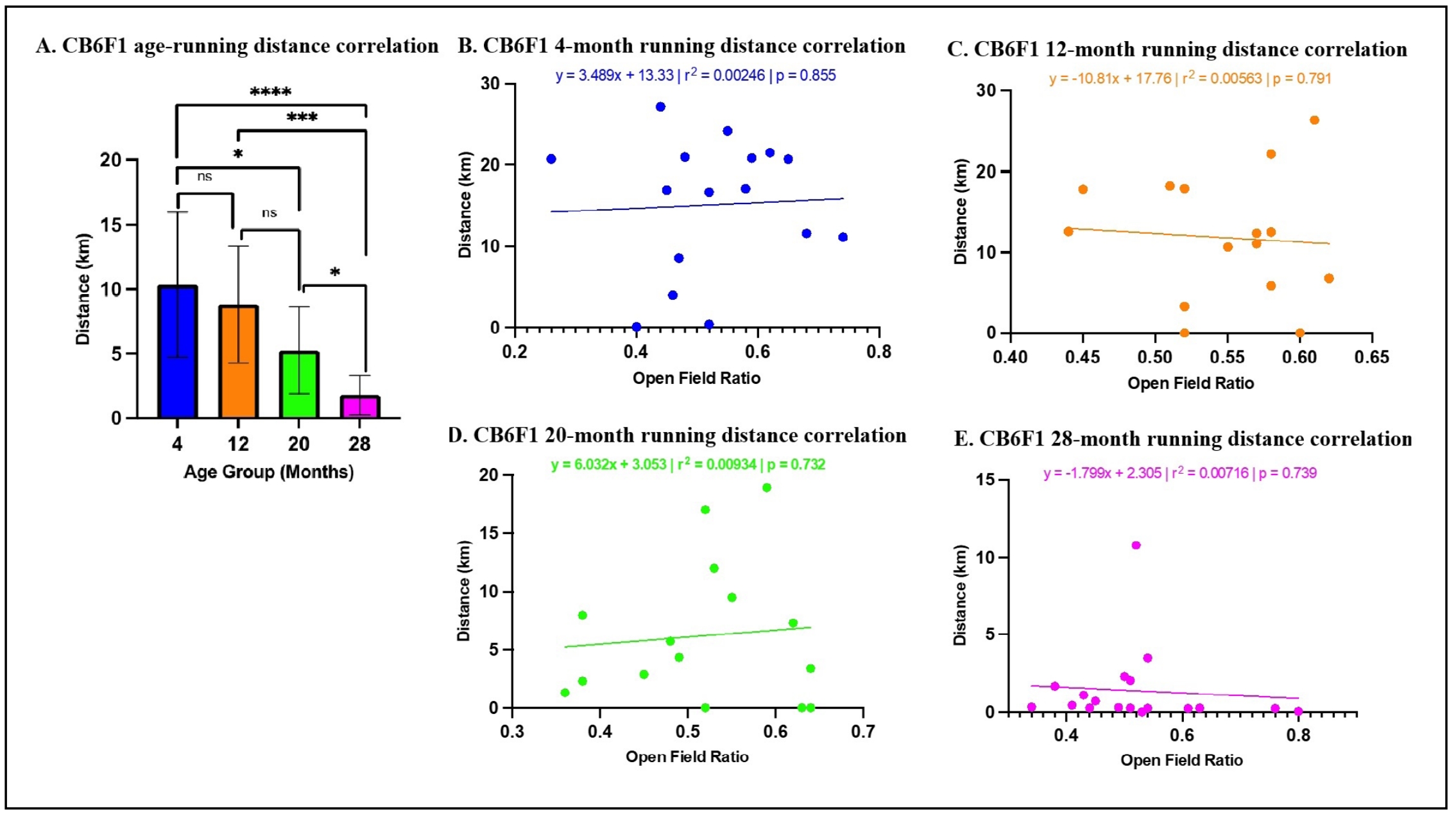
Figure 6. Relationship between OFR and voluntary wheel running distance in CB6F1 mice. (A) A statistically significant association was observed between age and wheel running. (B-E) There were no significant correlations among CB6F1 mice across age-groups between time spent in peripheral areas and total running distance (*P < 0.05, ***P < 0.001, ****P < 0.0001, not significant if P > 0.05, N = 19-28/cohort).
For C57BL/6 mice in voluntary wheel running, multiple linear regression analysis revealed that age was a significant predictor of distance traveled (ß= -0.375, P < 0.0001), with post-hoc analyses indicating that older CB6F1 mice exhibited a significant reduction in self-motivated running distances (Figure 7A). OFR was a significant predictor of distance traveled in the 28-month-old cohort (Figure 7E) but was not a significant predictor of distance traveled (ß= 3.52, P = 0.669) for 4-, 12-, and 20-month cohorts (Figure 7B-D). The overall model accounted for a significant proportion of the variance in distance traveled (R2 = 0.332, F (2,65) = 16.12, P < 0.0001).
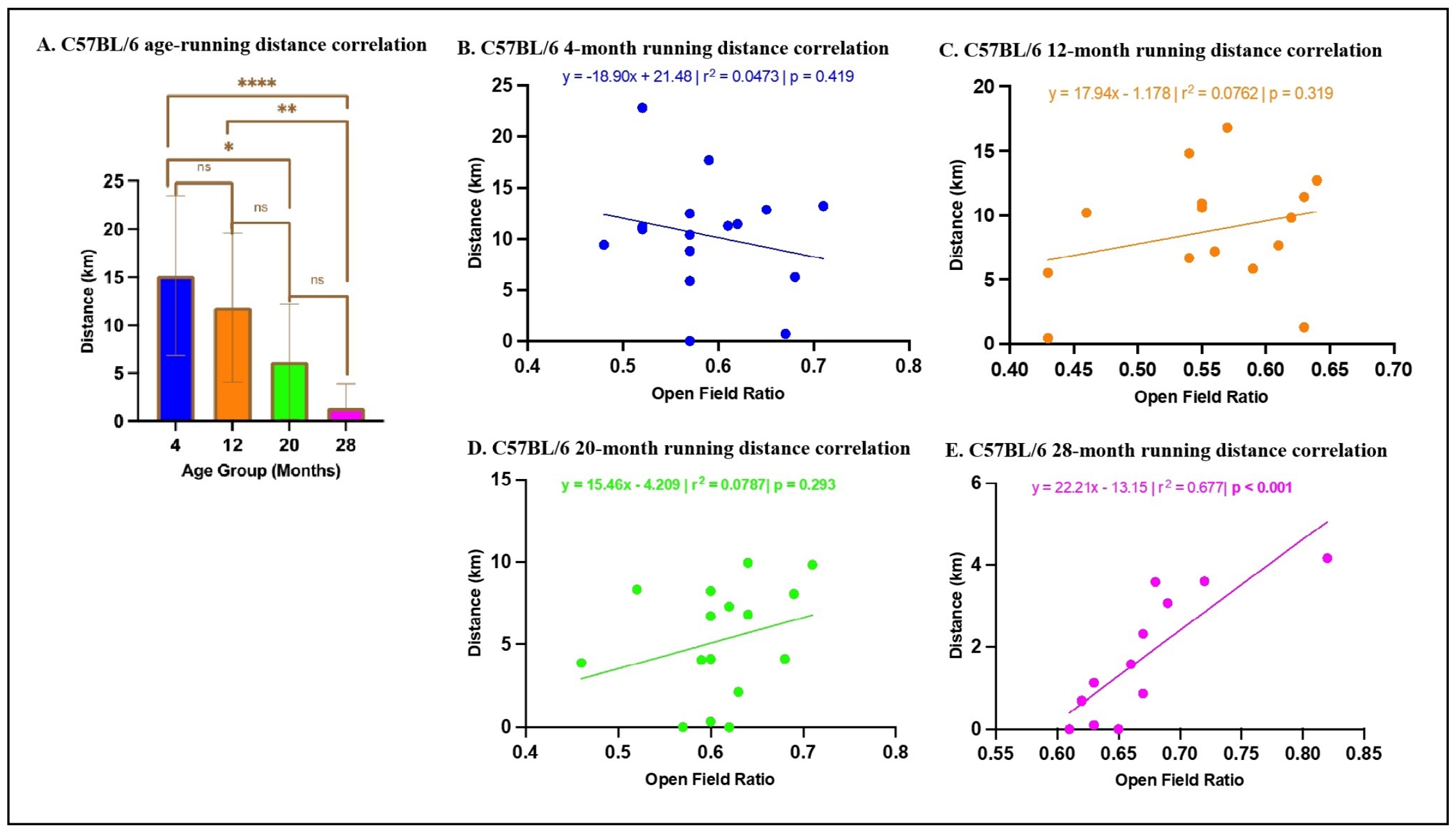
Figure 7. Relationship between OFR and voluntary wheel running distance in C57BL/6 mice. (A) A statistically significant association was observed between age and running distance. (B-D) There were no significant correlations among C57BL/6 mice 4-,12-, and 20-months old between time spent in peripheral areas and total running distance (*P < 0.05, **P < 0.01, ****P < 0.0001, not significant if P > 0.05, N = 19-28/cohort). (E) There was a significant correlation between time spent in peripheral areas and total running distance in 28-month-old C57BL/6 mice (P < 0.001, N= 12).
For grip strength, multiple linear regression analysis revealed that age was a significant predictor of grip strength in CB6F1 (ß= -1.240, P < 0.0001) and C57BL/6 mice (ß= -1.434, P < 0.0001), indicating that older mice in both strains exhibited a significant reduction in grip strength (Figure 8A & B). However, OFR was not a significant predictor of grip strength in either strain (CB6F1: ß= 4.230, P = 0.7830, C57BL/6: ß= -24.19, P = 0.3078). The overall model accounted for a significant proportion of the variance in grip strength for both strains (CB6F1: R2 = 0.3890, F (2,81) = 25.79, P < 0.0001, C57BL/6: R2 = 0.4525, F (2,78) = 32.24, P < 0.0001).
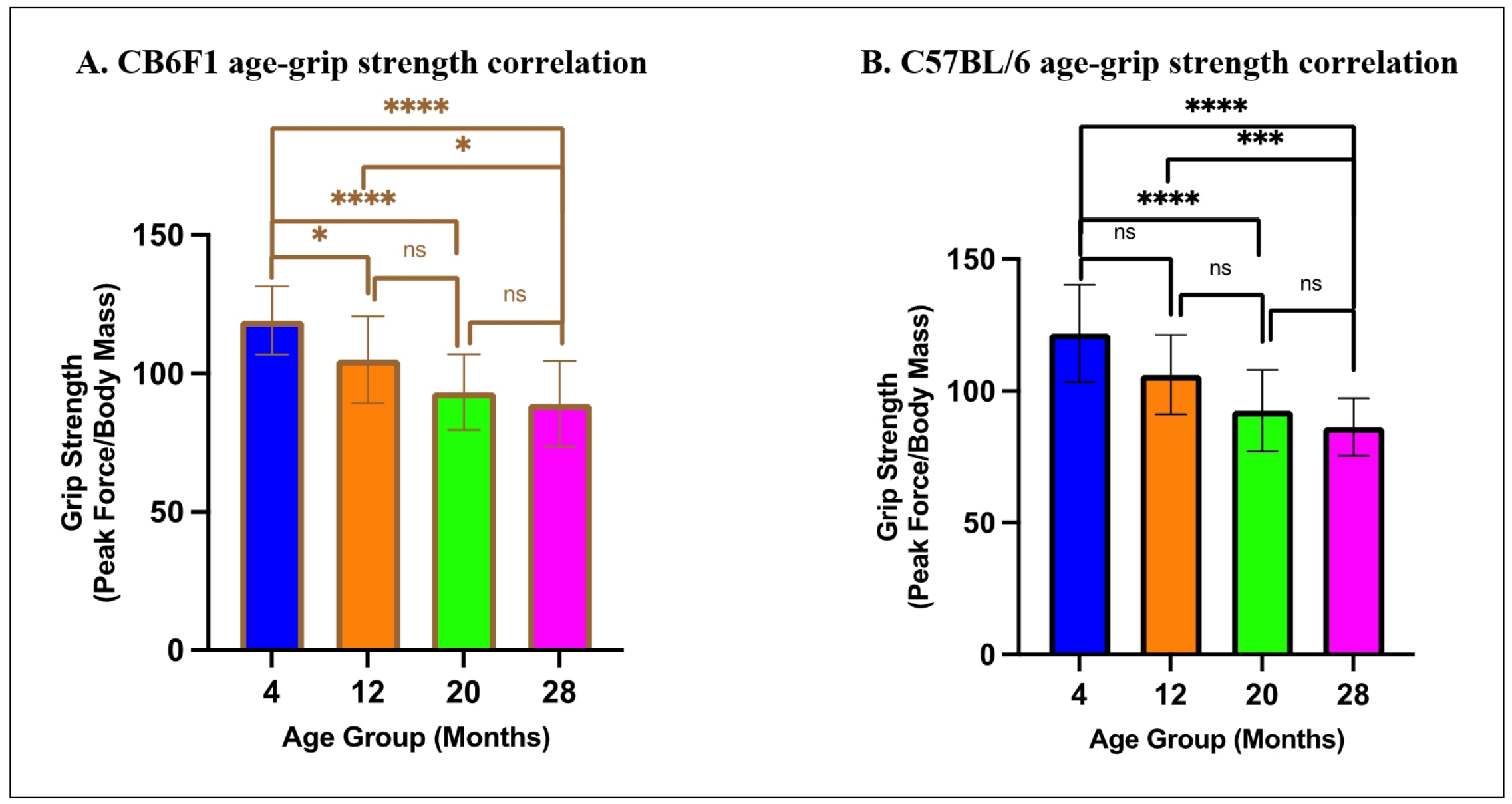
Figure 8. Relationship between age and grip strength. (A-B) There were significant correlations between age and grip strength in CB6F1 and C57BL/6 mice (*P < 0.05, ***P < 0.001, *****P < 0.0001, ns = not significant (P > 0.05), N = 19-27/cohort).
For rotarod performance, multiple linear regression analysis revealed that age was a significant predictor of rotarod performance in both CB6F1 (ß= -2.376, P < 0.0001) and C57BL/6 mice (ß= -1.997, P < 0.0001), indicating that older mice exhibited a significant reduction in rotarod performance (Figure 9A & B). However, OFR was not a significant predictor of rotarod performance in either strain (CB6F1: ß= 50.82, P = 0.1244, C57BL/6: ß= 47.94, P = 0.1244). The overall model accounted for a significant proportion of the variance in rotarod performance for both strains (CB6F1: R2 = 0.352, F (2,83) = 22.54, P < 0.0001, C57BL/6: R2 = 0.294, F (2,78) = 16.24, P < 0.0001).
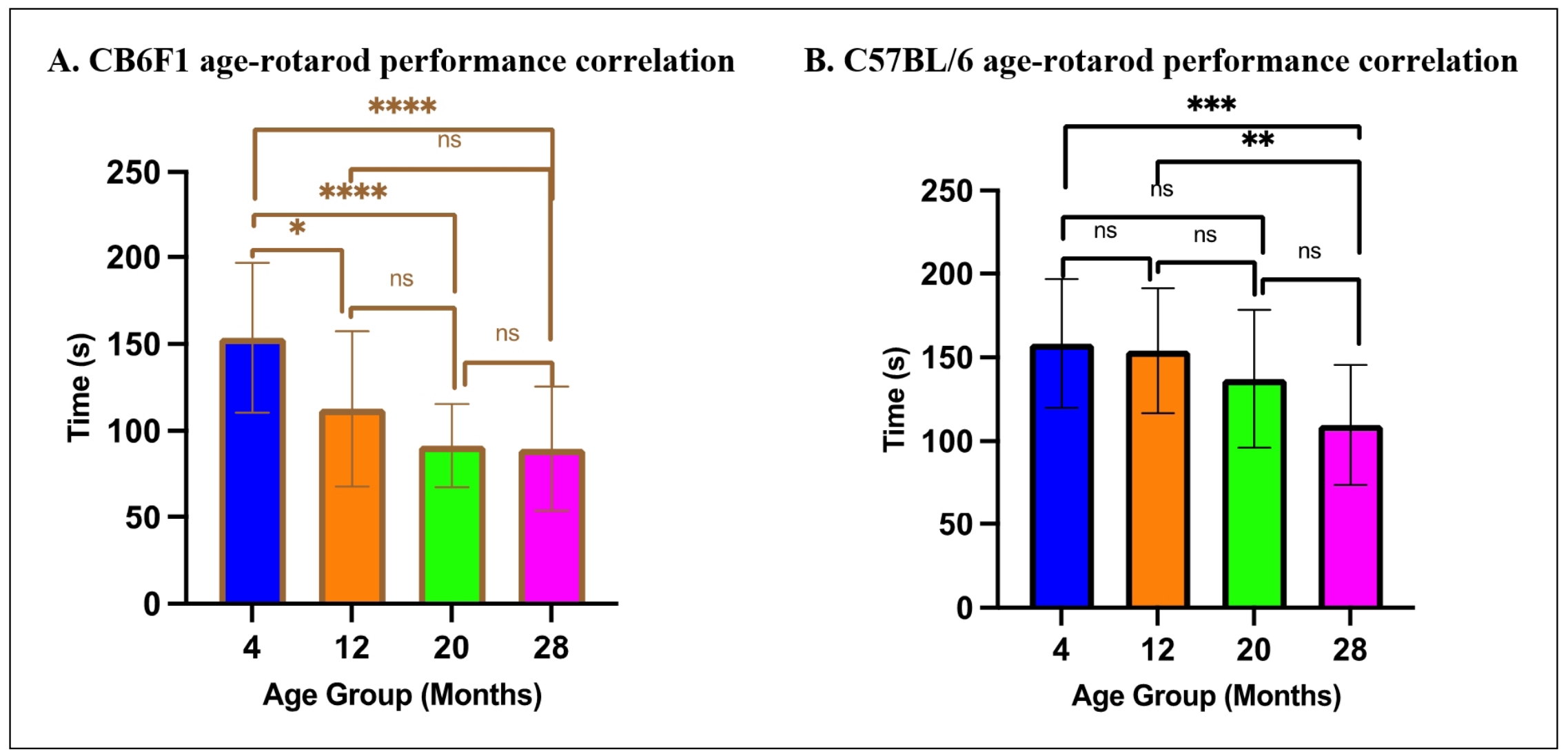
Figure 9. Relationship between age and rotarod performance. (A-B) There were significant correlations between age and rotarod performance in CB6F1 and C57BL/6 mice (*P < 0.05, ***P < 0.01, ***P < 0.001, ****P < 0.0001, ns = not significant (P > 0.05), N =19-29/cohort).
Discussion
Analysis of anxiety-related behaviors in the open-field photobeam testing system in relation to cognitive and performance
functions revealed insights into the relationships between anxiety, aging, and behavior in mice. Total movement, area region
preference, rearing time, and memory were all age-dependent in CB6F1 mice, while no correlation between the OFR and memory as well
as general motor function (as measured by the voluntary wheel running task, grip strength, and rotarod performance) were observed
in either strain. The OFT is useful to measure anxiety in mice based on thigmotaxis, the natural instinct of rodents to avoid open
spaces and especially with high intensity light due to their vulnerability to predation
[27]. This well-documented behavioral manifestation of anxiety is rooted in evolutionary
survival mechanisms and is associated with difficulties in emotional and spatial learning
[28]. The amount of time spent in peripheral regions emerged as a critical parameter of
anxiety. Older mice across strains exhibited heightened anxiety when exposed to a novel environment, as shown by increased time
spent in peripheral areas and time spent rearing. However, mice that displayed elevated anxiety levels did not demonstrate
worse performance in cognitive tasks, suggesting that anxiety and cognition operate independently of each other. A negative
correlation was also revealed between anxiety levels and distance traveled in the voluntary wheel running assessment, but only
in 28-month-old C57BL/6 mice. The impact of anxiety on learning and memory was not apparent in this study as we did not observe
an association of peripheral region preference and decreased long-term memory. However, previous research has emphasized the
role of neuronal circuits and neurotransmitter systems in anxiety-related cognitive impairment
[29, 30]. Stress hormones such as cortisol and
corticosterone are released during anxiety states and are known to impact the functioning of brain regions involved in memory
processing, such as the hippocampus and prefrontal cortex [31-36].
Furthermore, the rate of adult neurogenesis (AN), the concept of neuronal production in selective brain regions well after the
typical brain morphogenesis associated with neonatal development, has also been shown to independently affect cognition and
anxiety-like behaviors in rodents [37-39].
As the ventral dentate gyrus (DG) of the hippocampus is a well-documented area of AN in rodents
[40] and has been associated with anxiety-related behaviors and cognitive function
[27, 41-43],
further investigation of these variables would be highly relevant for future studies to elucidate the relationship between
anxiety and cognition.
Interestingly, while neither strain had statistically significant associations between memory performance and peripheral area
preference, CB6F1 mice demonstrated an age-related decrease in long-term memory while C57BL/6 mice did not, and associations
between anxiety-related behaviors and age were overall stronger. As C57BL/6 mice are often considered to have a lower baseline
level of anxiety without clearly understood reasons [44,
45], this observation suggests that it would be beneficial to conduct research in
elucidating differences and similarities in behavior and aging between C57BL/6 and CB6F1 mice.
A reduction in running distance in the voluntary wheelrunning assessment among mice with increased anxiety can be attributed to
reduced exploratory behavior and a diminished willingness to engage in physical activity
[46]. Such cautiousness, reluctance to take risks, and decreased engagement in
high-energy activities aligns with anxietyinduced behaviors, although a statistically significant association was only observed in
28-month-old C57BL/6 mice. Furthermore, no significant correlations with anxiety scores were found in either strain for the
rotarod or grip strength tests. The absence of a significant correlation between anxiety and motor function across ages and
strains, as assessed by the voluntary wheel running, rotarod, and grip strength tests, may be attributed to several factors.
Performance on these tests primarily reflects motor coordination and physical strength, while anxiety primarily affects emotional
and cognitive domains, with evidence from previous studies showing that cognitive changes may operate independently of gross motor
function [47]. While these tests may not directly elicit anxietyspecific responses in
mice, previous studies have found positive correlations between AN and exercise without affecting anxiety-related behavior in
rodents [48]. As the open-field test was performed within a month after the voluntary
wheel running and rotarod performance tasks and a few days prior to the grip strength assessment, the chronological order of
behavioral-assays may also have impacted our findings between behavior and cognition. Individual variability in mouse responses to
anxiety and motor function capabilities could also contribute to the observed absence of significant correlation. Further
investigations employing more specific anxiety-related assays or a broader array of behavioral tests may provide a more
comprehensive understanding of the relationship between anxiety and motor function in aging mice.
Conclusions
The results of this study highlight the influence of age and strain on anxiety responses in male mice, emphasizing the need for careful consideration of these factors when interpreting open-field test results. The observed preference for peripheral areas in novel environments suggests that physiological anxiety plays a role in mouse behavior and its implications for cognitive function. This study contributes to future research aimed at better understanding how anxiety and aging interact in male mice. It is inherent to conduct similar studies in female mice to determine and identify any sex-dependent differences, thus providing the basis for the development of new strategies to delay aging through anxiety management.
Declarations
Financial support and sponsorship
This project was supported by NIH grant R01 AG057381 (Ladiges, PI).
Conflicts of interest statement
No conflicts of interest.
Ethical approval
The study was approved by the University of Washington IACUC.
Conflicts of interest
Warren Ladiges is a member of the editorial board of Aging Pathobiology and Therapeutics. The authors declare that they have no conflicts and were not involved in the journal's review or decision regarding this manuscript.
References
1. Miceli M, & Castelfranchi C: Expectancy and emotion: OUP Oxford; 2014.
2. Rosen JB, & Schulkin J. From normal fear to pathological anxiety. Psychol Rev, 1998, 105(2): 325-350. [Crossref]
3. Rosen JB, Hamerman E, Sitcoske M, Glowa JR, & Schulkin J. Hyperexcitability: exaggerated fear-potentiated startle produced by partial amygdala kindling. Behav Neurosci, 1996, 110(1): 43-50. [Crossref]
4. Robinson OJ, Pike AC, Cornwell B, & Grillon C. The translational neural circuitry of anxiety. J Neurol Neurosurg Psychiatry, 2019, 90(12): 1353-1360. [Crossref]
5. Julian LJ. Measures of anxiety: state-trait anxiety inventory (STAI), beck anxiety inventory (BAI), and hospital anxiety and depression scale-anxiety (HADS-A). Arthritis Care Res, 2011, 63 (11): S467-472. [Crossref]
6. Shankman SA, Nelson BD, Sarapas C, Robison-Andrew EJ, Campbell ML, Altman SE, et al. A psychophysiological investigation of threat and reward sensitivity in individuals with panic disorder and/or major depressive disorder. J Abnorm Psychol, 2013, 122(2): 322-338. [Crossref]
7. Mohlman J, & Gorman JM. The role of executive functioning in CBT: a pilot study with anxious older adults. Behav Res Ther, 2005, 43(4): 447-465. [Crossref]
8. Hovenkamp-Hermelink JHM, Jeronimus BF, Myroniuk S, Riese H, & Schoevers RA. Predictors of persistence of anxiety disorders across the lifespan: a systematic review. Lancet Psychiatry, 2021, 8(5): 428-443. [Crossref]
9. Evans DL, Foa EB, Gur RE, Hendin H, O'Brien CP, Seligman ME, et al: Treating and preventing adolescent mental health disorders: what we know and what we don't know: Oxford University Press; 2005.
10. Sinoff G, & Werner P. Anxiety disorder and accompanying subjective memory loss in the elderly as a predictor of future cognitive decline. Int J Geriatr Psychiatry, 2003, 18(10): 951-959. [Crossref]
11. Kassem AM, Ganguli M, Yaffe K, Hanlon JT, Lopez OL, Wilson JW, et al. Anxiety symptoms and risk of cognitive decline in older community-dwelling men. Int Psychogeriatr, 2017, 29(7): 1137-1145. [Crossref]
12. Shackman AJ, Sarinopoulos I, Maxwell JS, Pizzagalli DA, Lavric A, & Davidson RJ. Anxiety selectively disrupts visuospatial working memory. Emotion, 2006, 6(1): 40-61. [Crossref]
13. Tan XJ, Dai YB, Wu WF, Warner M, & Gustafsson J. Anxiety in liver X receptor β knockout female mice with loss of glutamic acid decarboxylase in ventromedial prefrontal cortex. Proc Natl Acad Sci USA, 2012, 109(19): 7493- 7498. [Crossref]
14. Becker E, Orellana Rios CL, Lahmann C, Rücker G, Bauer J, & Boeker M. Anxiety as a risk factor of Alzheimer's disease and vascular dementia. Br J Psychiatry, 2018, 213(5): 654-660. [Crossref]
15. Roberts R, & Knopman DS. Classification and epidemiology of MCI. Clin Geriatr Med, 2013, 29(4): 753-772. [Crossref]
16. Beaudreau SA, & O'Hara R. Late-life anxiety and cognitive impairment: a review. Am J Geriatr Psychiatry, 2008, 16(10): 790-803. [Crossref]
17. Lyketoso C, Steele C, & Steinberg M: Behavioural disturbances in dementia. Reichel's care of the elderly-clinical aspects of ageing: Williams and Wilkins; 1999.
18. Steimer T. The biology of fear- and anxiety-related behaviors. Dialogues Clin Neurosci, 2002, 4(3): 231-249. [Crossref]
19. Kraeuter AK, Guest PC, & Sarnyai Z. The open field test for measuring locomotor activity and anxiety-like behavior. Methods Mol Biol, 2019, 1916: 99-103. [Crossref]
20. Ge X, Ciol MA, Pettan-Brewer C, Goh J, Rabinovitch P, & Ladiges W. Self-motivated and stress-response performance assays in mice are age-dependent. Exp Gerontol, 2017, 91: 1-4. [Crossref]
21. Lee WJ, Liao YC, Wang YF, Lin IF, Wang SJ, & Fuh JL. Plasma MCP-1 and cognitive decline in patients with Alzheimer's disease and mild cognitive impairment: a twoyear follow-up study. Sci Rep, 2018, 8(1): 1280-1290. [Crossref]
22. Lezak KR, Missig G, & Carlezon WA, Jr. Behavioral methods to study anxiety in rodents. Dialogues Clin Neurosci, 2017, 19(2): 181-191. [Crossref]
23. Daneshjoo S, Park JY, Moreno J, Rosenfeld M, Darvas M, & Ladiges W. A mouse model of naturally occurring agerelated cognitive impairment. Aging Pathobiol Ther, 2022, 4(3): 87-89. [Crossref]
24. Pettan-Brewer C, Touch DV, Wiley JC, Hopkins HC, Rabinovitch PS, & Ladiges WC. A novel radial water tread maze tracks age-related cognitive decline in mice. Pathobiol Aging Age Relat Dis, 2013, 3: 1-10. [Crossref]
25. Ge X, Cho A, Ciol MA, Pettan-Brewer C, Snyder J, Rabinovitch P, et al. Grip strength is potentially an early indicator of age-related decline in mice. Pathobiol Aging Age Relat Dis, 2016, 6: 32981. [Crossref]
26. Goh J, & Ladiges W. Voluntary wheel running in mice. Current protocols in mouse biology, 2015, 5(4): 283-290.
27. Bach DR. Cross-species anxiety tests in psychiatry: pitfalls and promises. Mol Psychiatry, 2022, 27(1): 154-163. [Crossref]
28. Küçük A, Gölgeli A, Saraymen R, & Koç N. Effects of age and anxiety on learning and memory. Behav Brain Res, 2008, 195(1): 147-152. [Crossref]
29. Chen GH, Wang C, Yangcheng HY, Liu RY, & Zhou JN. Agerelated changes in anxiety are task-specific in the senescence-accelerated prone mouse 8. Physiol Behav, 2007, 91(5): 644-651. [Crossref]
30. Abel T, Martin KC, Bartsch D, & Kandel ER. Memory suppressor genes: inhibitory constraints on the storage of long-term memory. Science, 1998, 279(5349): 338-341. [Crossref]
31. Kellett J, & Kokkinidis L. Extinction deficit and fear reinstatement after electrical stimulation of the amygdala: implications for kindling-associated fear and anxiety. Neuroscience, 2004, 127(2): 277-287. [Crossref]
32. Merz CJ, & Wolf OT. How stress hormones shape memories of fear and anxiety in humans. Neurosci Biobehav Rev, 2022, 142: 104901. [Crossref]
33. Anwar MJ, Pillai KK, Samad A, & Vohora D. Effect of escitalopram on cardiomyopathy-induced anxiety in mice. Hum Exp Toxicol, 2013, 32(6): 632-639. [Crossref]
34. Joung JY, Cho JH, Kim YH, Choi SH, & Son CG. A literature review for the mechanisms of stress-induced liver injury. Brain Behav, 2019, 9(3): e01235. [Crossref]
35. Oakley RH, Ren R, Cruz-Topete D, Bird GS, Myers PH, Boyle MC, et al. Essential role of stress hormone signaling in cardiomyocytes for the prevention of heart disease. Proc Natl Acad Sci USA, 2013, 110(42): 17035- 17040. [Crossref]
36. Zhang DY, & Anderson AS. The sympathetic nervous system and heart failure. Cardiology clinics, 2014, 32(1): 33- 45.
37. Revest JM, Dupret D, Koehl M, Funk-Reiter C, Grosjean N, Piazza PV, et al. Adult hippocampal neurogenesis is involved in anxiety-related behaviors. Mol Psychiatry, 2009, 14(10): 959-967. [Crossref]
38. Li YD, Luo YJ, Chen ZK, Quintanilla L, Cherasse Y, Zhang L, et al. Hypothalamic modulation of adult hippocampal neurogenesis in mice confers activity-dependent regulation of memory and anxiety-like behavior. Nat Neurosci, 2022, 25(5): 630-645. [Crossref]
39. Hill AS, Sahay A, & Hen R. Increasing adult hippocampal neurogenesis is sufficient to reduce anxiety and depression-like behaviors. Neuropsychopharmacology, 2015, 40(10): 2368-2378. [Crossref]
40. Kempermann G, Jessberger S, Steiner B, & Kronenberg G. Milestones of neuronal development in the adult hippocampus. Trends Neurosci, 2004, 27(8): 447-452. [Crossref]
41. Bannerman DM, Grubb M, Deacon RM, Yee BK, Feldon J, & Rawlins JN. Ventral hippocampal lesions affect anxiety but not spatial learning. Behav Brain Res, 2003, 139(1- 2): 197-213. [Crossref]
42. Weeden CS, Roberts JM, Kamm AM, & Kesner RP. The role of the ventral dentate gyrus in anxiety-based behaviors. Neurobiol Learn Mem, 2015, 118: 143-149. [Crossref]
43. Kheirbek MA, Drew LJ, Burghardt NS, Costantini DO, Tannenholz L, Ahmari SE, et al. Differential control of learning and anxiety along the dorsoventral axis of the dentate gyrus. Neuron, 2013, 77(5): 955-968. [Crossref]
44. An XL, Zou JX, Wu RY, Yang Y, Tai FD, Zeng SY, et al. Strain and sex differences in anxiety-like and social behaviors in C57BL/6J and BALB/cJ mice. Exp Anim, 2011, 60(2): 111-123. [Crossref]
45. Eltokhi A, Kurpiers B, & Pitzer C. Behavioral tests assessing neuropsychiatric phenotypes in adolescent mice reveal strain- and sex-specific effects. Sci Rep, 2020, 10(1): 11263. [Crossref]
46. McCartan CJ, Yap J, Firth J, Stubbs B, Tully MA, Best P, et al. Factors that influence participation in physical activity for anxiety or depression: a synthesis of qualitative evidence. Cochrane database of systematic reviews, 2020, 3: CD013547. [Crossref]
47. Nolte ED, Nolte KA, & Yan SS. Anxiety and task performance changes in an aging mouse model. Biochem Biophys Res Commun, 2019, 514(1): 246-251. [Crossref]
48. Garrett L, Lie DC, Hrabé de Angelis M, Wurst W, & Hölter SM. Voluntary wheel running in mice increases the rate of neurogenesis without affecting anxiety-related behaviour in single tests. BMC Neurosci, 2012, 13: 61-71. [Crossref]